БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 14. МЕТАБОЛІЗМ ЛІПІДІВ
14.4.Біосинтез вищих жирних кислот
Жирні кислоти синтезуються в організмі тварин переважно в абсорбтивний період, головним чином у клітинах печінці й жирової тканини. У цей період клітини зазвичай повністю забезпечені необхідними для синтезу жирних кислот de novo метаболічним субстратом, енергією та відновними еквівалентами у вигляді ацетил-КоА, АТФ і НАДФН відповідно.
Ферментні системи синтезу жирних кислот локалізовані в цитоплазмі й мітохондріальному матриксі, але цитоплазматична система синтезу значно переважає за активністю мітохондріаль- ну й тому біосинтез жирних кислот здійснюється головним чином у цитоплазмі.
Субстрат синтезу - молекули ацетил-КоА, що утворюються в мітохондріях при окисненні пірувату й жирних кислот, переносяться в цитоплазму за допомогою декількох транспортних систем: у формі ацетату, цитрату, а також за участю карнітину.
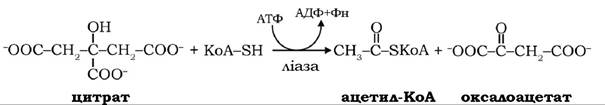
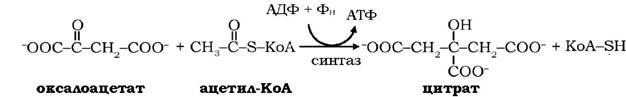
При гальмуванні циклу трикарбонових кислот надлишком АТФ цитрат, який утворюється в реакції між ацетил-КоА і оксалоацетатом за участю цитратсинтази, переноситься в цитоплазму за допомогою трикарбоксилаттранспортуючої системи:
У цитоплазмі цитрат розщеплюється АТФ-цитратліазою, утворюючи цитозольний ацетил-КоА:
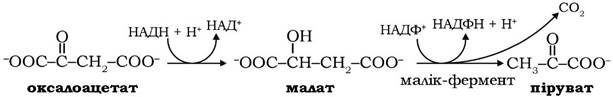
Оксалоацетат, один із продуктів цитратліазної реакції, перетворюється на піруват, який повертається в мітохондрії:
Молекули НАДФ+, які відновлюються в цій реакції за участю малатдегідрогенази (малік-ферменту), використовуються як відновні еквіваленти у відновних реакціях біосинтезу жирних кислот.
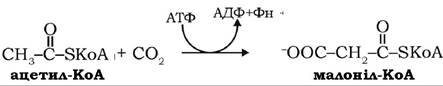
Ацетил-КоА, утворений у цитратліазній реакції, карбоксилюється цитозольною ацетил-КоА-карбоксилазою з утворенням безпосереднього попередника синтезу жирних кислот - малоніл-КоА:
Біотиновий фермент ацетил-КоА-карбоксилаза є регуляторним ферментом, він регулює швидкість усього процесу біосинтезу жирних кислот. Його алостеричним модулятором служить цитрат, тобто в даному випадку цитрат виконує дві важливі функції: як переносник ацетил-КоА і як активатор ацетил-КоА-карбоксилази.
Наступні реакції цього біосинтетичного шляху здійснюються безпосередньо мультиферментною системою, яка називається синтазною системою для жирних кислот., або пальмітатсинтазою. Цей ферментний комплекс, загальна молекулярна маса якого становить ~ 400 кДа, об'єднує сім ферментів, котрі беруть участь у біосинтезі пальмітинової кислоти (рис. 14.5). Центральним компонентом цієї системи є ацилпереносний білок (АПБ), який функціонує в процесі синтезу як молекулярний кронштейн, що послідовно переносить проміжні метаболіти синтезу від одного ферменту синтази до наступного. Простетична група АПБ - 4'-фосфопантетеїн (похідне вітаміну пантотенової кислоти) у процесі побудови вуглецевого ланцюга жирної кислоти утворює ефірні зв'язки з кожним проміжним продуктом, аналогічно процесу, який відбувається в піруватдегідрогеназному ферментному комплексі (див. розд. 12, п. 12.1.3).
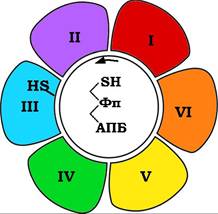
Рис. 14.5. Схема пальмітатсинтазного ферментного комплексу:
I - ацетилтрансфераза; II - малонілтрансфераза; III - 3-кетоацил-АПБ-синтаза;
IV - 3-кетоацил-АПБ-редуктаза; V - 3-гідроксіацил-АПБ-дегідратаза; VI - еноїл-АПБ-редуктаза
Синтазна система для жирних кислот каталізує сумарну реакцію побудови 16-вуглецевого ланцюга молекули пальмітинової кислоти з однієї молекули ацетил-КоА і семи молекул малоніл-КоА:
![]()
У цьому синтезі молекула ацетил-КоА слугує матрицею, до якої послідовно приєднуються двокарбонові фрагменти від малоніл- КоА. Нарощування вуглецевого ланцюга молекули ВЖК йде від її метильної групи та оксогрупи (положення 16 і 15 відповідно) у напрямку карбоксильної групи пальмітату.
Починається синтез пальмітату за участю ферментів ацетил- трансферази й малонілтрансферази, які каталізують приєднання субстратів синтезу - ацетильної та малонільної груп до двох SH-груп синтази. Далі побудова жирної кислоти здійснюється циклічно в чотири етапи (рис. 14.6).
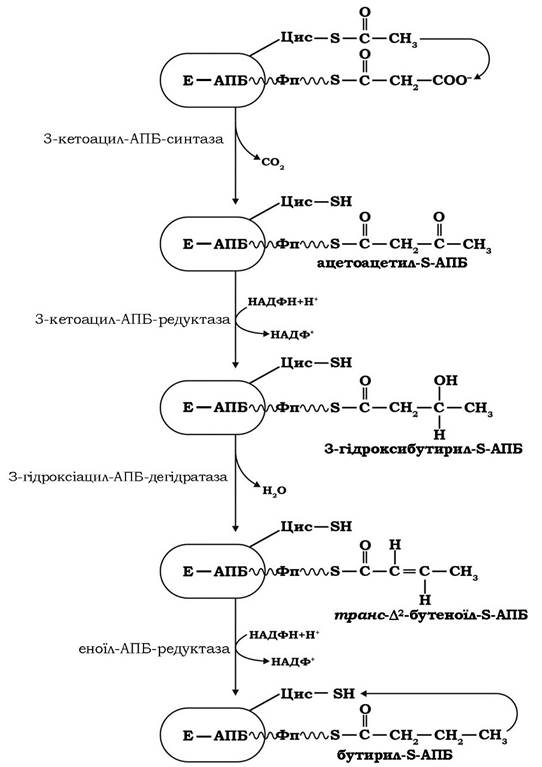
Рис. 14.6. Послідовність реакцій одного циклу синтезу пальмітату
Спочатку ацетильна група за дії 3-кетоацил-АПБ-синтази переноситься з БН-групи цистеїну синтази на малонільну групу, приєднану до фосфопантетеїну АПБ. У реакції конденсації між цими двома групами вивільняється СО2, який попередньо був включений у молекулу малоніл-КоА в ацетил-КоА-карбоксилазній реакції.
Потім ацетоацетил-Б-АПБ відновлюється 3-кетоацил-АПБ-редуктазою за участю НАДФН у 3-гідроксибутирил-S-АПБ. Далі має місце реакція дегідратації, що каталізується 3-гідроксіацил-АПБ- дегідратазою з утворенням трансД2-бутеноїл-S-АПБ.
На завершальному етапі першого циклу реакцій синтезу відбувається відновлення подвійного зв'язку транс∆2-бутеноїл-S- АПБ за участю еноїл-АПБ-редуктази й бутирильна група від бутирил-S-АПБ переноситься на вільну SH-групу цистеїну синтази - на місце, яке початково займала ацетильна група.
Далі відбувається новий цикл реакцій подовження вуглецевого ланцюга ще на один двокарбоновий фрагмент, тобто цикли повторюються ще сім разів і в результаті утворюється кінцевий продукт - пальмітоїл-S-АПБ. На цьому процес синтезу жирної кислоти закінчується. За дії гідролази молекула пальмітинової (С16) кислоти відщеплюється від АПБ синтази.
Слід відмітити, що синтез однієї молекули пальмітату потребує енергії семи макроергічних зв'язків АТФ для утворення тіоефір- них зв'язків у молекулах субстратів ацетил-КоА і малоніл-КоА і відновного потенціалу 14 молекул НАДФН для насичення подвійних зв'язків.
Молекули НАДФН для редуктазних реакцій синтезу надходять від двох джерел: від реакцій окиснення глюкозо-6-фосфату на пентозофосфатному шляху та окиснення малату в малатдегідрогеназній реакції.
Синтазна система для жирних кислот не спроможна синтезувати жирні кислоти з числом атомів вуглецю більше 16. Але подальше нарощування вуглецевого ланцюга пальмітинової кислоти можливо за участю ферментних систем ендоплазматичного ретикулума шляхом приєднання ацетильних груп від малоніл-КоА або в мітохондріях, де приєднуються ацетильні групи від молекул ацетил-КоА.
Пальмітинова і стеаринова кислоти є попередниками для двох моноєнових жирних кислот пальмітолеїнової (С16∆^9) і олеїнової (С18∆9), які утворюються за участю ацил-КоА-оксигенази та молекулярного кисню і НАДФН:
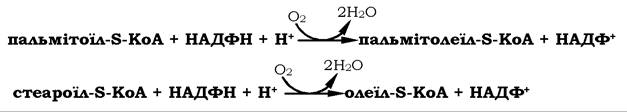
У тканинах тварин (ссавців) не синтезуються лінолева (С18∆9, 12) і α-ліноленова (С18∆9,12,15) кислоти, які є попередниками двох інших важливих поліненасичених кислот - γ-лінолевої (С18∆6,9,12) і арахідонової (С20∆5,8,11,14) - незамінних попередників біологічно активних сполук - ейкозаноїдів.
Регуляція біосинтезу вищих жирних кислот у тканинах тварин здійснюється молекулярними механізмами, що діють як на клітинному рівні, так і рівні організму в цілому (рис. 14.7).
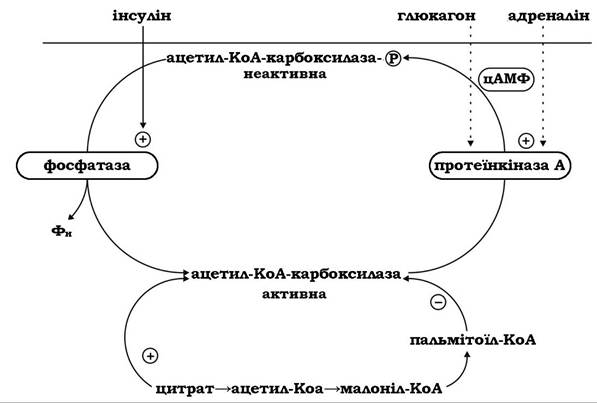
Рис. 14.7. Схема регуляції активності ацетил-КоА-карбоксилази
На клітинному рівні функціонують механізми алостеричної регуляції та ковалентної модифікації ключових ферментів біосинтезу ВЖК. Активність головного регуляторного ферменту цього синтетичного шляху - ацетил-КоА-карбоксилази, що визначає швидкість синтезу ВЖК, контролюється його алостеричними модуляторами: фермент активується цитратом та інгібується пальмітоїл-КоА.
Пальмітоїл-КоА є також інгібітором пальмітоїл-синтетази за механізмом негативного зворотного зв’язку, або ретроінгібування.
В абсорбтивний період під дією гормону інсуліну активується фосфатаза, яка дефосфорилює неактивну ацетил-КоА-карбоксилазу, переводячи її в активний стан. У постабсорбтивний період, а також при стресі, фізичному навантаженні тощо, гормони глюкагон, адреналін та інші, що діють через аденілатциклазну систему, інактивують ацетил-КоА-карбоксилазу шляхом фосфорилювання її активною протеїнкіназою А.
Синтез жирних кислот регулюється також дієтою. Вуглеводна дієта приводить до індукції ряду ферментів вуглеводного й ліпідного обмінів, які каталізують перетворення проміжних метаболітів розщеплення глюкози на жирні кислоти.
Малоніл-КоА, який утворюється в ацетил-КоА-карбоксилазній реакції, інгібує також активність карнітинацилтрансферази, гальмуючи тим самим надходження та окиснення жирних кислот у мітохондріях. Зниження рівня малоніл-КоА в цитозолі й пов'язана з цим активація карнітинацилтрансферази стимулює перехід жирних кислот із цитоплазми в мітохондрії та їхнє β-окиснення.