БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 14. МЕТАБОЛІЗМ ЛІПІДІВ
14.5.Синтез і розщеплення жирів у тканинах
Жирні кислоти, що синтезуються de novo або потрапляють у тканини тварин у результаті гідролізу триацилгліцеролів ліпопротеїнів крові (хіломікрони і ліпопротеїни дуже низької густини), не накопичуються в тканинах у вільному стані, тому що швидко перетворюються на компактну й зручну для зберігання форму - жири.
Найактивніше утворення жирів відбувається в абсорбтивний період, в умовах високої концентрації глюкози в крові, печінці, жировій та інших тканинах.
Попередниками триацилгліцеролів слугують активовані форми гліцеролу та вищих жирних кислот. Активна форма гліцеролу - гліцерол-3-фосфат може утворюватися двома шляхами: фосфорилюванням вільного гліцеролу або гідруванням дигідроксіацето- нфосфату. У клітинах печінки, нирок, слизової оболонки кишечнику, де присутня активна гліцеролкіназа, гліцерол фосфорилюється γ-фосфатною групою АТФ:
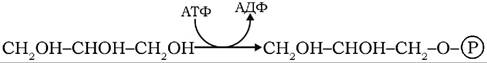
У клітинах жирової тканини, м'язах і печінці за участю гліцеролфосфатдегідрогенази гліцерол-3-фосфат утворюється відновленням дигідроксіацетонфосфату:
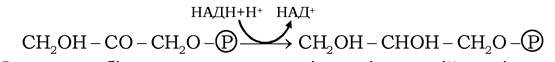
Ферменти біосинтезу триацилгліцеролів асоційовані в комплекс триацилгліцеролсинтази, локалізований на мембранах ендоплазматичного ретикулума.
Активація жирних кислот, котрі залучаються до синтезу ліпідів, каталізується ацил-КоА-синтазами:
![]()
Активовані жирні кислоти у вигляді КоА-ефірів у наступних реакціях переносяться на гліцерол-3-фосфат гліцеролфосфат- ацилтрансферазою:
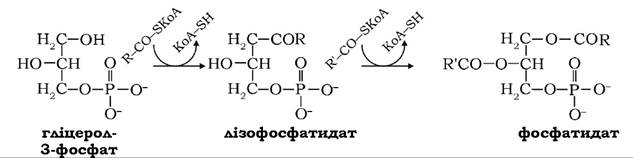
Продукт реакції - діацилгліцерол-3-фосфат, або фосфатидна кислота, утворюється через проміжний метаболіт - лізофосфатидат.
У ході синтезу триацилгліцеролів фосфатидат дефосфорилюється за дії фосфатидатфосфатази з утворенням 1,2-діацил- гліцеролу, котрий у реакції ацилювання, що каталізується діацил- гліцеролаиилтрансферазою, перетворюється в триацилгліцерол:
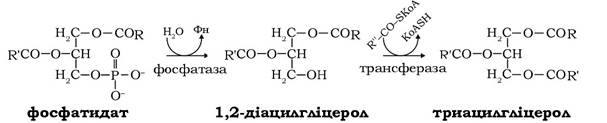
Найінтенсивніше жири синтезуються в печінці та жировій тканини. Триацилгліцероли, що синтезуються в печінці, в основному використовуються в інших тканинах, до яких вонитранспортуються у складі ліпопротеїнів плазми крові. Триацил- гліцероли, що утворюються в жировій тканині, накопичуються в цитоплазмі адипоцитів у вигляді крапель. Частка резервованих триацилгліцеролів становить приблизно 65 % маси всієї тканини. Депоновані таким чином жири за звичайних умов життєдіяльності організмів і достатньої калорійності раціону є довгостроковим резервом метаболічної енергії, оскільки для нагальних потреб у крові постійно перебувають ліпопротеїни, які виконують функцію безпосередніх постачальників жирних кислот до периферійних тканин. За таких умов в організмі дорослих тварин і людини встановлюється динамічна рівновага між інтенсивністю процесів біосинтезу й розщеплення жиру в жирових депо. Жирова тканина характеризується активним обміном речовин, тому при відносно сталому вмісті жирів резерви триацилгліцеролів постійно обмінюються й повне оновлення жирів здійснюється за декілька днів.
При зростанні калорійності їжі, переїданні або при зменшенні енергетичних затрат (гіподинамії) надлишок калорій іде на синтез жирів незалежно від характеру дієти (вуглеводної, білкової чи змішаної). У разі підвищених енергетичних витрат, тривалого голодування, стресових навантажень тощо вміст резервованих у жировій тканині триацилгліцеролів зменшується внаслідок тривалої та не компенсованої біосинтезом мобілізації жиру в жировій тканині.
Мобілізацією жиру називають процес гідролізу (ліполізу) триацилгліцеролів, що каталізується внутрішньоклітинною ліпазою. Активність ліпази жирової тканини контролюється гормонами (рис. 14.8). Гормони - АКТГ, адреналін, норадреналін, глюкагон - стимулюють її активність через аденілатциклазну систему шляхом підвищення рівня цАМФ. Активується ліполіз також тиреоїдними гормонами, які гальмують активність ферменту фосфодіестерази циклонуклеотидів, підтримуючи таким чином високий рівень цАМФ у клітині. цАМФ активує цАМФ-залежну протеїнкіназу, що фосфорилює неактивну ліпазу і перетворює її в активну ліпазу а. У результаті ліполізу жиру, який каталізується гормоночутливою ліпазою, утворюється гліцерол і вільні жирні кислоти, котрі вивільняються з адипоцитів у кров:
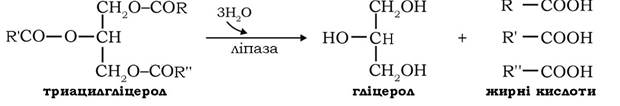
Гліцерол використовується печінкою як субстрат глюконеогенезу, а жирні кислоти транспортуються альбумінами крові, що служать у даному випадку переносниками жирних кислот із жирової тканини в інші периферійні тканини, де жирні кислоти можуть бути використані як енергетичний субстрат. Інсулін і простагландин Е блокують гормональну стимуляцію аденілатииклази, гальмуючи тим самим ліполіз.
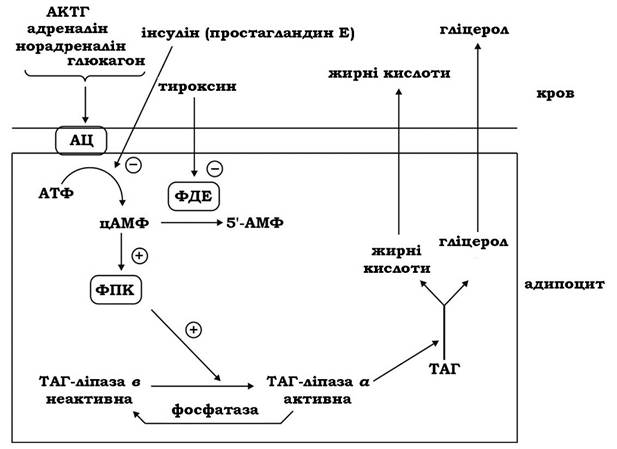
Рис. 14.8. Гормональна регуляція мобілізації жиру з жирової тканини:
АЦ - аденілатциклаза; ФДЕ - фосфодіестераза циклонуклеотидів;
ФПК - фосфопротеїнкіназа; ТАГ — триацилгліцероли
Гормоночутлива цАМФ-залежна ліпаза триацилгліцеролів є ліпопротеїном, який складається з білка (48 %), фосфоліпідів (45 %) і холестеролу (6 %). В активному центрі ферменту міститься залишок серину, тому ліпаза, як і інші "серинові" ферменти, інгібується фторидом натрію і фосфорорганічними сполуками.