БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
26. ЭВОЛЮЦИЯ, ИЛИ ИСТОРИЯ ЖИЗНИ НА ЗЕМЛЕ
26.7. Данные в пользу теории эволюции
26.7.7. Сравнительная эмбриология
Фон Бэр (1792—1867), изучая эмбриональное развитие у представителей разных групп позвоночных, обнаружил удивительное структурное сходство во всех этих группах, особенно на стадиях дробления, гаструляции и ранних стадиях дифференцировки. Геккель (1834—1919) высказал мысль, что это сходство имеет эволюционное значение. Он сформулировал закон рекапитуляции, согласно которому «онтогенез повторяет филогенез», т. е. стадии, через которые проходит организм в процессе своего развития, повторяют эволюционную историю той группы, к которой он относится. Хотя этот принцип слишком упрощает истинное положение дел, он привлекателен и в известной степени справедлив. Изучая одни только зародышевые стадии любой группы позвоночных, невозможно определить, к какой группе они относятся.
Как можно видеть на рис. 26.15, только на относительно поздней стадии развития зародыш начинает приобретать некоторое сходство с соответствующей взрослой особью. На определенных сравнимых стадиях развития у зародышей всех позвоночных имеются следующие структуры.
1. Карманообразные впячивания эктодермы и растущие им навстречу выпячивания стенок глотки. У рыб они соединяются и срастаются, а затем прорываются и образуют настоящие жаберные щели, обеспечивающие газообмен. У других групп позвоночных сохраняется одна такая щель, из которой развивается евстахиева труба и слуховой проход.
2. Сегментированные миотомы (зачатки скелетной мускулатуры), которые можно видеть в хвостообразной структуре. Они сохраняются только у некоторых видов.
3. Один круг кровообращения с двухкамерным сердцем, не разделенным на правую и левую половины; во взрослом состоянии такое строение сердца сохраняется только у рыб.
По мере развития зародыша позвоночного у него происходят изменения, в результате которых он приобретает признаки рыбы, амфибии, рептилии, птицы или млекопитающего, в зависимости от того, к какой группе относились его родители. Изначальное сходство между зародышами объясняют тем, что все они, а следовательно, и классы, к которым они относятся, имели общего предка. Если у организмов в процессе эмбрионального развития образуются зачатки, из которых не будут развиваться функционирующие органы, то их можно интерпретировать лишь как остатки структур, имевшихся у его предков. Закон рекапитуляции, однако, не может быть принят безоговорочно, так как ни у одного из ныне живущих организмов нельзя обнаружить все признаки его предполагаемых предков. Но представляется вероятным, что организмы сохраняют механизмы развития, унаследованные от своих предков. Поэтому возможно, что на разных стадиях эмбриогенеза у данного организма будут наблюдаться черты структурного сходства с зародышами предковых форм. Последующие адаптации к иным условиям среды и образу жизни изменят дальнейший ход развития. Как показывают наблюдения, чем ближе группы, к которым относят два данных организма на основании общих гомологичных дефинитивных структур, тем дольше сохраняется их сходство на эмбриональных стадиях. Организмы, адаптированные к определенному образу жизни и определенному местообитанию, не типичным для той крупной группы, к которой они принадлежат, менее сходны с другими членами этой группы и в процессе эмбрионального развития. Это видно на примере развития двух паразитических плоских червей Fasciola и Taenia: у них имеется ряд личиночных стадий с адаптациями к вторичным хозяевам, отсутствующим в жизненном цикле свободноживущих плоских червей Planaria. Точно так же у дождевого червя Lumbricus, ведущего наземный образ жизни, нет стадии трохофоры — личинки, типичной для более примитивных кольчатых червей. На этих примерах ясно видна ограниченность геккелевского закона рекапитуляции.
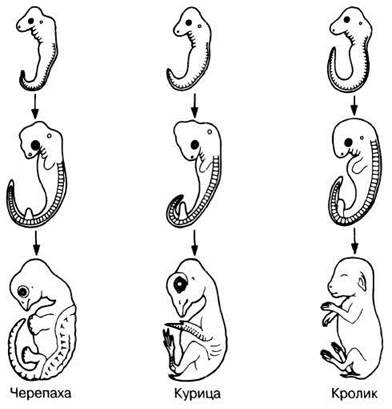
Рис. 26.15. Сравнение стадий эмбрионального развития на примере представителей трех классов позвоночных.
Изучение эмбриогенеза представителей крупных групп организмов выявляет черты структурного сходства, очевидные на эмбриональных и личиночных стадиях, но отсутствующие у взрослых особей. Эти наблюдения интерпретируются как признаки филогенетического родства между различными группами организмов, что подразумевает существование эволюционного процесса. По характеру дробления зиготы и судьбе бластопора организмы с трехслойным зародышем можно разделить на две группы — первичноротых и вторичноротых. Для первичноротых характерно спиральное дробление, а бластопор превращается в ротовое отверстие взрослой особи. Такой тип развития свойствен кольчатым червям, моллюскам и членистоногим. Для вторичноротыххарактерно радиальное дробление, и бластопор превращается у них в анальное отверстие взрослой особи. Так происходит развитие иглокожих и хордовых. Различия между этими двумя группами показаны на рис. 26.16. Именно такого рода данные помогли выяснить вопрос о филогенетических родственных связях иглокожих. Строение взрослых иглокожих заставляет рассматривать их как один из типов беспозвоночных, но эмбриональное развитие, протекающее у них по типу, характерному для вторичноротых, подтверждает их близость к линии развития хордовых. Этот пример показывает, что вопрос о филогенетических связях нельзя решать на основе одних лишь сведений о гомологичных структурах взрослых особей.
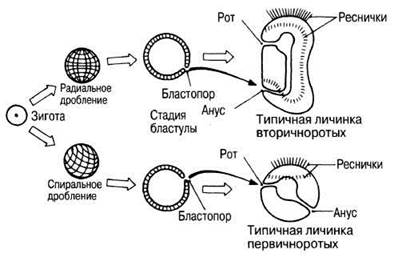
Рис. 26.16. Ранние стадии развития вторичноротых и первичноротых.
В результате изучения эмбриогенеза растений также получены данные, свидетельствующие о прогрессивной эволюции различных групп, однако изученные примеры не так хорошо документированы, как в царстве животных. У мхов и папоротников гаметофит, на ранних стадиях развития представленный протонемой, образующийся в результате прорастания спор, по своей структуре, физиологии и типу роста сходен с нитчатыми зелеными водорослями; поэтому принято считать, что и мхи, и папоротники происходят от этих водорослей. Чередование поколений в жизненных циклах растений и его различные варианты (отражающие адаптацию к разным условиям среды) можно интерпретировать как дополнительные доводы в пользу существования эволюционных связей между различными группами растений.
Голосеменные — это группа, промежуточная по своим признакам между формами, приспособленными к жизни на суше, и формами, у которых встреча гамет непременно должна происходить в воде. У саговников мужской гаметофит похож на легкую сухую микроспору (пыльцевое зерно) покрытосеменных тем, что он тоже распространяется ветром. В процессе развития мужского гаметофита одна из его клеток образует пыльцевую трубку, как у покрытосеменных, однако вместо того чтобы доставлять неподвижную мужскую гамету к архегонию, в пыльцевой трубке образуются два спермия, снабженные жгутиками, которые подплывают к семязачатку и осуществляют оплодотворение. Поэтому можно предположить, что саговники — группа, промежуточная между низшими растениями и покрытосеменными, а это в свою очередь позволяет думать, что в царстве растений существует филогенетическая непрерывность.
Существование группы организмов, обладающих чертами, общими для двух других групп, находящихся на разных уровнях сложности или адаптированных к различным местообитаниям, можно интерпретировать как указание на филогенетическую непрерывность между тремя этими группами — на происхождение одной группы (такой, как покрытосеменные) от другой (такой, как низшие растения) через промежуточную группу (саговники). Многие из таких промежуточных форм вымерли, и только изучая палеонтологическую летопись, можно выявить эволюционную последовательность. Во многих случаях промежуточные формы найти не удается. Эти формы соответствуют «недостающим звеньям» (разрывам) в палеонтологической летописи. Возможно также, что эти звенья и не существуют, что соответствует гипотезе прерывистого равновесия, описанной в разд. 26.7.1. Однако отсутствие таких звеньев можно объяснить тем, что их остатки либо не сохранились, либо еще не найдены. Промежуточным звеном между кольчатыми червями и членистоногими служит группа Onychopliora, представленная родом Peripatus, который обладает признаками как кольчатых червей, так и членистоногих (рис. 26.17). С кольчатыми червями перипатуса сближают строение стенки тела, в которой имеются кольцевые и продольные мышцы, нечленистые конечности, напоминающие параподии, выделительные канальцы, сходные с нефридиями, имеющиеся в южном сегменте, и двойная брюшная нервная цепочка. К признакам сходства с членистоногими относятся хитиновая кутикула, трахеи, дыхальца и незамкнутая система кровообращения. Другая группа «живых ископаемых», служащая промежуточным звеном между рыбами и амфибиями, представлена двоякодышащими.
Хотя большая часть этих данных указывает на существование той или иной формы эволюционного процесса, следует помнить, что твердыми данными об этом мы не располагаем.

Рис. 26.17. Примитивное членистоногое Peripalus.