БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
22. РОСТ И РАЗВИТИЕ
22.4. Рост и развитие цветковых растений
22.4.6. Латеральные меристемы и вторичный рост
После первичного роста происходит вторичный рост как результат активности латеральных меристем. Вторичный рост приводит к утолщению стебля или корня. Он обычно связан с отложением больших количеств вторичной ксилемы — древесины, которая совершенно изменяет первичную структуру и составляет характерную особенность деревьев и кустарников.
Известно два типа латеральной меристемы — сосудистый камбий, из которого образуется новая проводящая ткань, и пробковый камбий, или феллоген, который развивается позднее, замещая эпидермис, разрывающийся при утолщении тела растения.
Сосудистый камбий
Этот камбий состоит из клеток двух типов — веретеновидных инициалей и лучевых инициалей (рис. 22.20). Веретеновидные инициали — это узкие вытянутые клетки, которые делятся митотически, образуя снаружи от себя вторичную флоэму, а вовнутрь — вторичную ксилему. Количество образующейся ксилемы обычно превышает количество флоэмы. Последовательные деления показаны на рис. 22.21. Вторичная флоэма содержит ситовидные трубки, клетки-спутницы, склеренхимные волокна, склереиды, а также паренхиму.
Лучевые инициали имеют почти сферическую форму и делятся митотически, давая начало паренхимным клеткам, которые накапливаются и образуют лучи между соседними флоэмой и ксилемой.
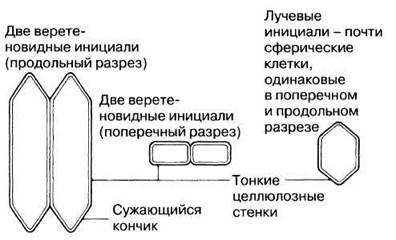
Рис. 22.20. Веретеновидные и лучевые инициали.
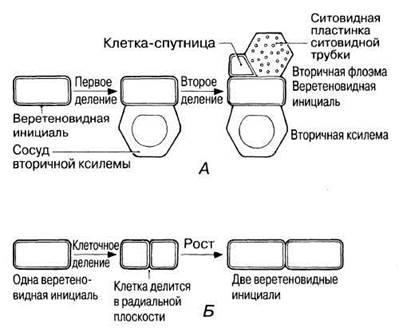
Рис. 22.21. А. Два последовательных деления веретеновидной инициали, в результате которых образуются ксилема и флоэма (в поперечном разрезе). На самом деле дифференцировка ксилемы и флоэмы до представленного здесь уровня занимает довольно много времени, в течение которого должно образоваться много клеток. Б. Деление веретеновидной инициали с образованием новой веретеновидной инициали (в поперечном разрезе).
Вторичный рост стебля у деревянистых двудольных растений
Сосудистый камбий первоначально располагается между первичной ксилемой и первичной флоэмой проводящих пучков; его образование из апикальной меристемы показано на рис. 22.17. Он становится активным почти сразу же после завершения первичной дифференцировки клеток. На рис. 22.22 показаны ранние стадии вторичного утолщения стебля типичного деревянистого двудольного растения.
На рис. 22.22, А представлена первичная структура стебля (перицикл не показан для простоты). А на рис. 22.22, Б можно видеть сплошной камбиальный цилиндр. На рис. 22.22, В изображено сплошное кольцо камбиального утолщения. Здесь веретеновидные инициали образовали большое количество вторичной ксилемы и меньшее количество вторичной флоэмы, а лучевые инициали — лучевую паренхиму. С утолщением стебля должна увеличиваться и длина окружности ствола. Это становится возможным благодаря делению камбиальных клеток в радиальном направлении (рис. 22.21).
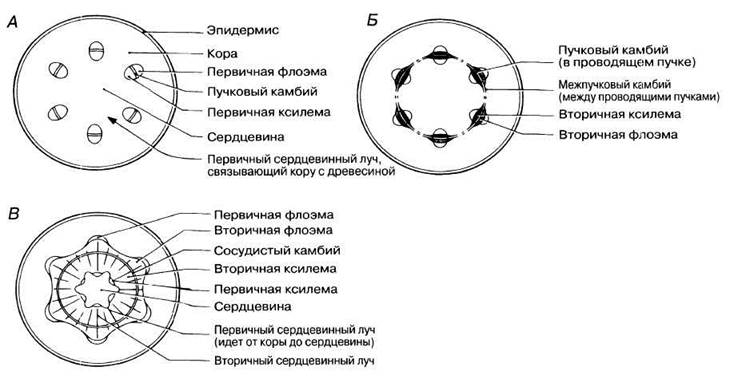
Рис. 22.22. Ранние стадии вторичного утолщения ствола у типичного деревянистого двудольного растения. А. Первичное строение стебля. Б. По мере того как сердцевинные лучи становятся меристематическими и растут наружу от проводящих пучков, камбий образует сплошной цилиндр. Тем временем происходит образование вторичной ксилемы и вторичной флоэмы из имеющегося камбия. В. Сплошное кольцо вторичного утолщения сформировалось. Утолщение особенно сильно выражено на местах первых проводящих пучков, где начиналась активность камбия.
Возникшие вначале лучевые инициали образуют первичные сердцевинные лучи, которые тянутся от сердцевины до коры, в отличие от вторичных сердцевинных лучей, образующихся из лучевых инициалей, возникших позднее. Эти лучи служат живым связующим звеном между сердцевиной и корой. Они обеспечивают радиальный перенос в стебле и воды, и минеральных солей из ксилемы и питательных веществ из флоэмы. Кроме того, через межклеточные пространства может происходить газообмен путем диффузии.
Лучи могут также служить местом для запасания питательных веществ, что имеет особенно важное значение в периоды покоя, например зимой. В трехмерном изображении они имеют вид продольных радиальных пластов, так как лучевые инициали лежат друг над другом, образуя радиальные колонки, как показано на рис. 22.23. На этом рисунке представлено строение древесины (вторичной ксилемы) и содержащиеся в ней лучи на поперечном тангенциальном и продольно-радиальном срезах.
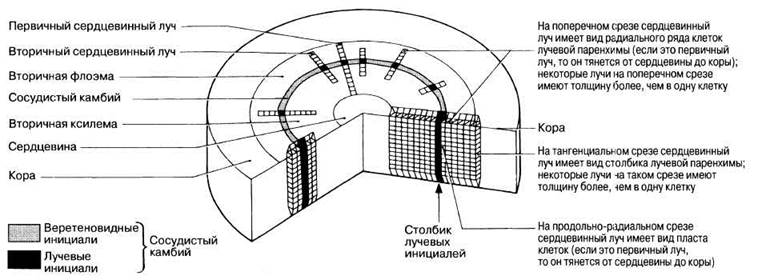
Рис. 22.23. Схематическое изображение первичных и вторичных сердцевинных лучей в стволе типичного деревянистого двудольного растения. Справа показан первичный, а слева — вторичный луч.
На рис. 22.24 изображена часть стебля деревянистого двудольного растения на третий год роста; видны большие количества образовавшейся вторичной ксилемы. На рис. 22.25 показаны срезы стволов липы (Тiliа) в возрасте трех и пяти лет.
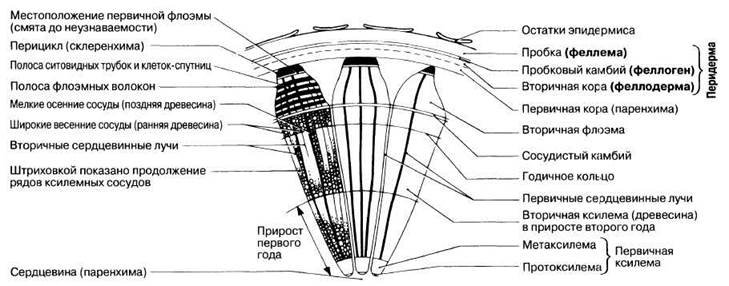
Рис. 22.24. Ствол типичного деревянистого двудольного растения — липы (Тiliа) — на третьем году роста (возраст два года) в поперечном разрезе. Детали строения вторичной флоэмы, вторичной ксилемы и вторичных сердцевинных лучей показаны только елевом секторе.
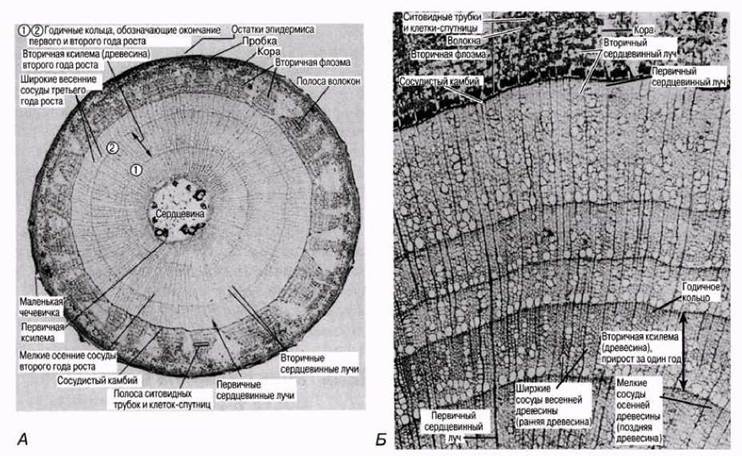
Рис. 22.25. А. Поперечный срез ветки Тiliа vulgaris в возрасте двух лет (третий год роста); х2,2. Б. Часть поперечного среза ветки Тiliа vulgaris в возрасте пяти лет (шестой год роста); x11,5.
Годичные кольца
В областях с умеренным климатом ежегодно весной рост возобновляется. Первыми при этом образуются широкие тонкостенные сосуды, способные проводить большие количества воды. Вода необходима для инициации роста, особенно для появления новых клеток, например в развивающихся листьях. Позднее сосудов образуется меньше, они становятся уже, а их стенки толще. В течение зимы камбий пребывает в состоянии покоя. Осенняя древесина, образующаяся в когте вегетационного периода, непосредственно перед прекращением роста, резко отличается от примыкающей к ней весенней древесины следующего года. В результате получаются так называемые годичные кольца, ясно различимые на рис. 22.25. Если сосуды сконцентрированы в ранней древесине, то древесину называют кольцепоровой; если же они распределены равномерно, так что годичные кольца выражены менее четко, то древесину называют рассеянно-поровой. В тропиках сезонные засухи могут вызвать сходные колебания активности камбия, Ширина годичного кольца варьирует, частично в зависимости от климата: благоприятный климат способствует образованию большего количества древесины, расстояния между кольцами бывают больше. Эта зависимость используется в двух областях науки — в дендроклиматологии и дендрохронологии. Дендроклиматология занимается изучением климата на основе данных о годичных кольцах, например выявляет корреляции между результатами метеорологических наблюдений и ростом деревьев в определенной местности или же исследует климатические события, происходившие несколько сотен или даже тысяч лет назад. Возраст самых древних из ныне живущих деревьев — остистых сосен — достигает примерно 5000 лет, а среди ископаемых остатков деревьев известны еще более древние.
Дендрохронология занимается датировкой древесины на основе изучения характера колец. Их рисунок служит своего рода «отпечатками пальцев», позволяя установить, в какое время росло дерево. Это дает возможность определить возраст бревен, найденных при археологических раскопках, возраст старых построек, кораблей и т. п.
Ядро и заболонь
По мере старения дерева древесина в центре ствола перестает выполнять проводящие функции и заполняется темными отложениями, такими как таннины. Эту внутреннюю часть называют ядром, или ядровой древесиной, а более влажную проводящую часть — заболонью.
Пробка и чечевички
По мере роста вторичной ксилемы кнаружи ткани, прилегающие к ней с этой стороны, все больше спрессовываются и одновременно растягиваются, потому что увеличивается и длина окружности ствола. Это оказывает влияние на эпидермис, кору, первичную флоэму и почти на всю вторичную флоэму, кроме только что образовавшегося слоя. Эпидермис в конце концов разрывается и замещается пробкой в результате активности второй латеральной меристемы — пробкового камбия, или феллогена; этот камбий обычно образуется прямо под эпидермисом. Пробка, или феллема, откладывается снаружи от пробкового камбия, тогда как с внутренней стороны образуются один или два слоя паренхимы. Эти слои неотличимы от первичной коры и образуют феллодерму, или вторичную кору. Феллоген, пробка и феллодерма в совокупности образуют перидерму (рис. 22.24).
При созревании пробковых клеток их стенки пропитываются жировым веществом — суберином, непроницаемым для воды и газов. Клетки постепенно отмирают, утрачивая живое содержимое, и наполняются либо воздухом, либо смолой или таннинами. Старые мертвые пробковые клетки смыкаются друг с другом и окружают ствол, предохраняя его от высыхания, инфекций и механических повреждений. По мере утолщения ствола эти клетки спрессовываются и могут в конце кондов заместиться лежащими под ними более молодыми клетками. Если бы пробковый слой был сплошным, обмен кислородом и диоксидом углерода между живыми клетками стебля и наружной средой был бы невозможен и эти клетки погибали бы. Однако в пробке имеются чечевички — щелевидные отверстия, расположенные на разных расстояниях друг от друга и заполненные рыхлой массой мертвых тонкостенных клеток, не содержащих суберина. Чечевички образуются пробковым камбием; у них имеются межклеточные воздушные полости, делающие возможным газообмен. Схема строения чечевички представлена на рис. 22.26.
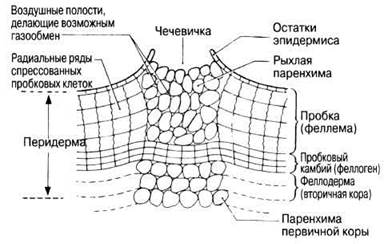
Рис. 22.26. Чечевичка в вертикальном разрезе (содержимое клеток не показано).
Кора
В конце концов деревянистый стебель покрывается слоем, известным под названием коры. Кора — нестрогий термин, которым обозначают либо все ткани, лежащие снаружи от проводящей системы, либо — в более узком смысле — ткани, лежащие снаружи от пробкового камбия. При сдирании коры с дерева обычно удаляются все ткани, вплоть до сосудистого камбия — тонкого слоя клеток, который легко разрывается.
Пробковый камбий обычно возобновляется каждый год. Часто пробковый камбий образуется из вторичной флоэмы; в таких случаях кора с течением времени становится слоистой, что обусловлено чередованием слоев вторичной флоэмы и коры.