БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
22. РОСТ И РАЗВИТИЕ
22.4. Рост и развитие цветковых растений
22.4.5. Первичный рост корня
Строение апикальной меристемы типичного корня представлено на рис. 22.19. На самом кончике апикальной меристемы находится покоящийся центр — группа инициалей (меристематических клеток), от которых в конечном счете происходят все другие клетки корня, но которые делятся значительно медленнее, чем их потомки в окружающей апикальной меристеме. Снаружи от них образуются клетки корневого чехлика; они превращаются в крупные паренхимные клетки, которые защищают апикальную меристему при углублении корня в почву. Эти клетки непрерывно снашиваются и замещаются новыми. Они несут, кроме того, важную дополнительную функцию — служат рецепторами гравитации: в них содержатся крупные крахмальные зерна, которые играют роль статолитов, опускаясь на «дно» клеток под действием силы тяжести. Подробнее роль этих зерен описана в разд. 16.2.2.
Позади покоящегося центра можно видеть правильные ряды клеток и зоны меристемы, уже описанные для побега, а именно протодерму, основную меристему и прокамбий (рис. 22.19). В корне прокамбием называют весь центральный цилиндр, хотя в зрелом состоянии он содержит непроводящие ткани перицикла и сердцевины, если она имеется.
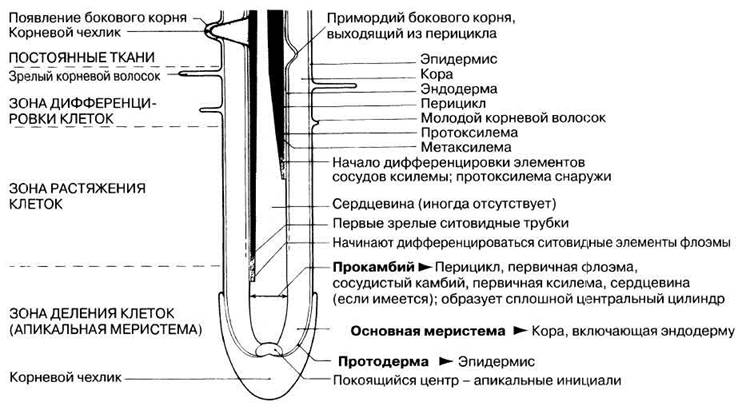
Рис. 22.19. Апикальная меристема типичного корня (в продольном разрезе). Справа показана дифференциация ксилемы, а слева — дифференциация флоэмы. На самом деле ксилема и флоэма чередуются, располагаясь на разных радиусах по окружности корня. Кроме того, в действительности зона растяжения в корне длиннее.
Зона клеточного деления обычно занимает 1—2 мм позади кончика корня и слегка перекрывается с зоной растяжения клеток. Кончики корней служат удобным материалом для изучения митоза, и один из применяемых для этого методов описан в разд. 23.3. Позади зоны деления рост происходит главным образом за счет растяжения клеток; клетки увеличиваются в размерах так же, как это описано для побега и показано на рис. 22.18. Зона увеличения размеров клеток находится на расстоянии примерно 10 мм от кончика корня и удлинение этих клеток проталкивает кончик корня вниз, заставляя его углубляться в почву.
Некоторая дифференцировка клеток начинается еще в зоне деления с развитием первых ситовидных элементов флоэмы (рис. 22.19). На продольных срезах кончика корня четко видны продольные ряды ситовидных элементов, достигающих все большей зрелости по мере удаления от кончика корня, пока они не превращаются во вполне сформировавшиеся ситовидные трубки. Развитие флоэмы идет в направлении снаружи внутрь, к центральной оси корня.
Еще дальше от кончика корня в зоне растяжения начинают дифференцироваться сосуды ксилемы, тоже центростремительно (экзархная ксилема) в отличие от центробежной дифференцировки в стебле (эндархная ксилема). Первыми образуются, как и в стебле, сосудистые элементы протоксилемы и у них происходит такая же лигнификация и растяжение по мере роста окружающих клеток. Затем их функции переходят к метаксилеме, которая развивается позднее и созревает в зоне дифференцировки после того, как прекратится растяжение клеток. Ксилема часто доходит до центральной оси корня, и в таких случаях сердцевина не образуется.
Развитие легче изучать в корнях, чем в побегах. В побегах тяжи прокамбия, идущие к листьям, осложняют распространение развивающихся тканей. Особенно хорошо наблюдать развитие ксилемы на давленых препаратах апикальных участков тонких корней, например проростков кресс-салата, при соответствующем окрашивании.
Дифференцировка заканчивается, когда все клетки перестают удлиняться. К этому времени завершается и развитие корневых волосков из эпидермиса.