БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
23. НЕПРЕРЫВНОСТЬ ЖИЗНИ
23.4. Мейоз
Мейоз (от греч. méiōsis — уменьшение) — форма деления ядра, сопровождающегося уменьшением числа хромосом от диплоидного (2n) до гаплоидного (n). Как и при митозе, при этом во время интерфазы происходит репликация ДНК в родительской клетке, однако за этим следуют два цикла делений ядра и делений клеток, известные как первое деление мейоза (мейоз I) и второе деление мейоза (мейоз II). Таким образом, одна диплоидная клетка дает начало четырем гаплоидным клеткам, как это схематически показано на рис. 23.10.
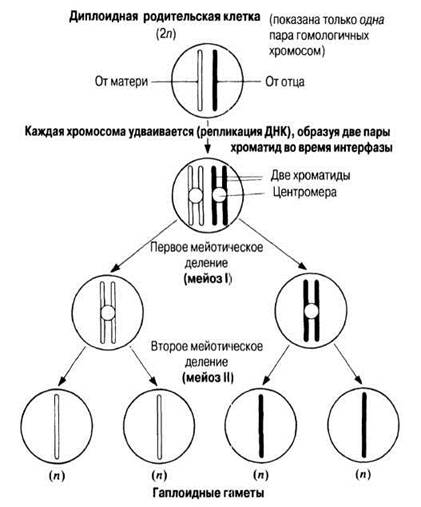
Рис. 23.10. Схематическое изображение основных этапов мейоза на примере дупликации одной хромосомы и двух последующих делений ядра и клеток. Обратите внимание, что, как и при митозе, хромосомы могут быть одиночными или двойными структурами. Две части удвоившейся хромосомы называют хроматидами.
Мейоз происходит при образовании спермиев и яйцеклеток (гаметогенез) у животных (гл. 21) и при образовании спор у растений.
Подобно митозу, мейоз — непрерывный процесс, но его тоже можно ради удобства подразделить на профазу, метафазу, анафазу и телофазу. Эти стадии происходят в первом делении мейоза и еще раз повторяются во втором. Поведение хромосом в течение этих стадий представлено на рис. 23.11, где показано деление ядра, содержащего четыре хромосомы (2n = 4), т. е. две пары гомологичных хромосом.
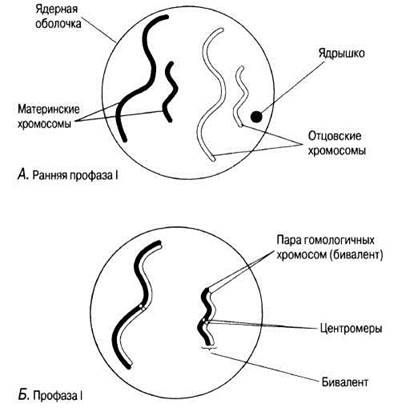
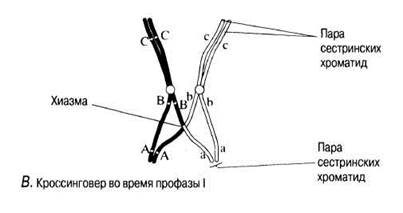
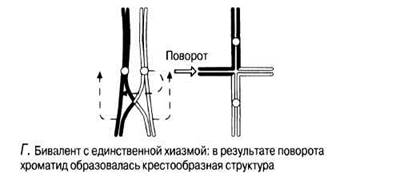
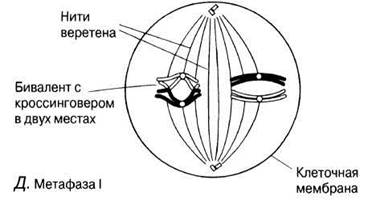
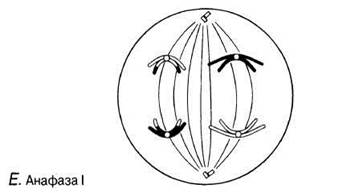
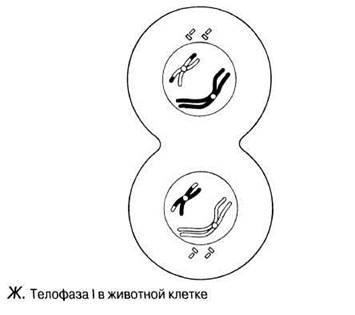
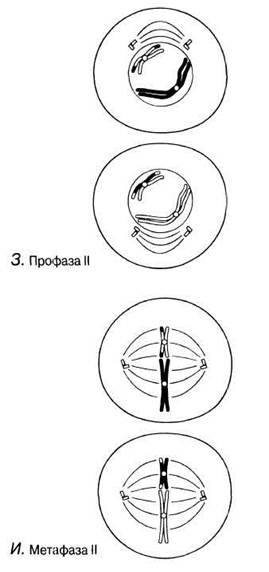
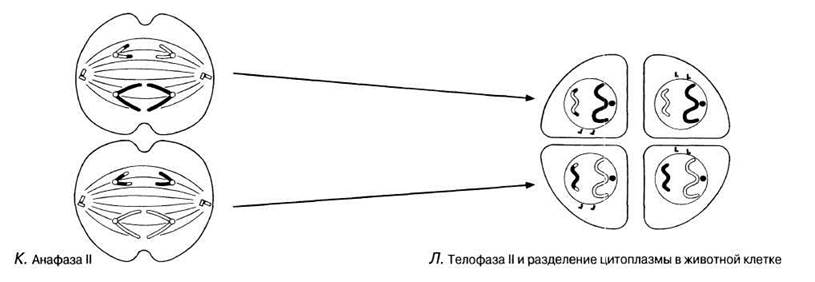
Рис. 23.11. А — Г. Мейоз в животной клетке. Д—Ж. Мейоз в животной клетке. 3 —Л. Мейоз в животной клетке.
МЕЙОЗ I
Фотографии мейоза представлены на рис. 23.12—23.14.
Профаза I
Самая продолжительная фаза.
А. Хромосомы укорачиваются и становятся видны как дискретные структуры.
Б. Гомологичные хромосомы спариваются. Этот процесс называется синапсисом, а каждая пара хромосом — бивалентом; одна хромосома происходит от родителя мужского пола, другая — от родителя женского пола. Обе хромосомы бивалента имеют одинаковую длину, их центромеры занимают одинаковое положение и они обычно состоят из одинакового числа генов, расположенных в одном и том же порядке. Биваленты укорачиваются и утолщаются, частично за счет скручивания. Хромосомы и центромеры на этой стадии четко различимы.
В. Гомологичные хромосомы как бы отталкиваются одна от другой и частично разделяются. Становится видно, что каждая их них состоит из двух хроматид. Хромосомы все еще соединены между собой в нескольких точках. Эти точки называются хиазмами (от греч. chiasma — перекрест). В каждой хиазме происходит обмен между хроматидами, осуществляющийся в результате разрыва и воссоединения между каждыми двумя из четырех нитей, имеющихся в каждой хиазме. В результате гены из одной хромосомы (например, отцовской А, В, С) оказываются связанными с генами из другой хромосомы (материнской а, b, с), что приводит к новым генным комбинациям в образующихся хроматидах. Этот процесс называют кроссинговером.
Г. Хроматиды гомологичных хромосом продолжают отталкиваться друг от друга,- и биваленты принимают определенную конфигурацию в зависимости от числа хиазм. (Биваленты с одной хиазмой образуют кресты, с двумя хиазмами принимают кольцевидную форму, а с тремя и более хиазмами образуют петли, расположенные под прямыми углами одни к другим.) К концу профазы I:
1) все хромосомы полностью сокращаются и интенсивно окрашиваются;
2) центриоли (если они имеются) мигрируют к полюсам;
3) ядрышки и ядерные оболочки дезинтегрированы;
4) образуются нити веретена.
Метафаза I
Биваленты выстраиваются у экватора веретена, прикрепляясь к нитям центромерами.
Анафаза I
Нити веретена тянут гомологичные хромосомы, начиная с центромер, к противоположным полюсам веретена. В результате хромосомы разделяются на два гаплоидных набора, по одному на каждом полюсе веретена.
Телофаза I
Расхождение гомологичных хромосом к противоположным полюсам соответствует окончанию мейоза I. Число хромосом уменьшилось вдвое, но они все еще состоят из двух хроматид каждая. Если произошел кроссинговер, то эти хроматиды генетически неидентичны и при втором мейотическом делении должны будут разойтись. Веретёна и их нити обычно исчезают. У животных и у некоторых растений хроматиды обычно раскручиваются, на каждом полюсе вновь образуется ядерная оболочка и ядро вступает в интерфазу. Затем происходит дробление (у животных) или формирование клеточной стенки (у растений) как при митозе. У многих растений не наблюдается ни телофазы, ни образования клеточной стенки, ни интерфазы, и клетка из анафазы I прямо переходит в профазу второго мейотического деления.
Интерфаза II
Эта стадия обычно имеется только у животных клеток. Ее продолжительность варьирует. Дальнейшей репликации ДНК не происходит.
МЕЙОЗ II
Мейоз II сходен с митозом.
Профаза II
В случае отсутствия интерфазы II эта стадия также отсутствует. Ядрышки и ядерные мембраны разрушаются, а хроматиды укорачиваются и утолщаются. Центриоли, если они имеются, перемещаются к противоположным полюсам клетки, и к концу профазы II появляются новые нити веретена. Они расположены под прямыми углами к веретену мейоза I.
Метафаза II
Хромосомы выстраиваются по отдельности вокруг экватора веретена.
Анафаза II
Центромеры делятся и нити веретена растаскивают их, а за ними и хроматиды к противоположным полюсам.
Телофаза II
Происходит так же, как телофаза митоза с той лишь разницей, что образуются четыре гаплоидные дочерние клетки. Хромосомы раскручиваются, удлиняются и становятся плохо различимыми. Нити веретена исчезают, а центриоли реплицируются. Вокруг каждого ядра вновь образуется ядерная оболочка, но ядро содержит теперь половину числа хромосом исходной родительской клетки (оно гаплоидно). При последующем дроблении (у животных) или образовании клеточной стенки (у растений) из единственной родительской клетки получается четыре дочерних клетки.
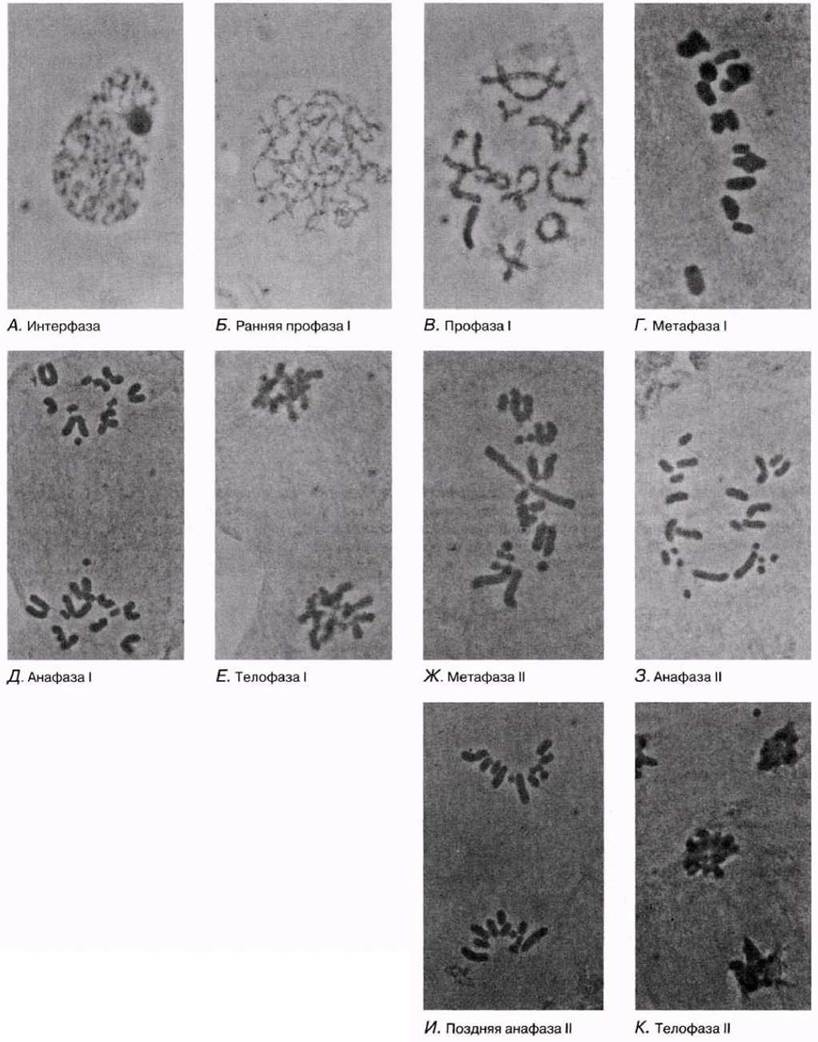
Рис. 23.12. Стадии мейоза в животной клетке. Показана также интерфаза.
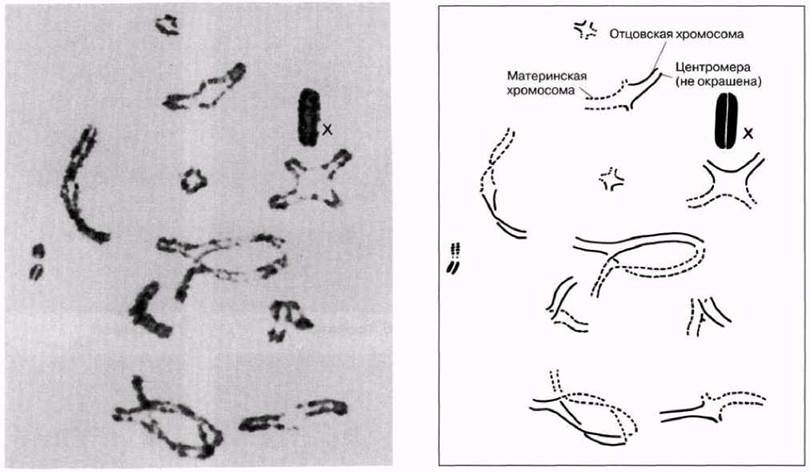
Рис. 23.13. Кроссинговер в профазе I в клетке странствующей саранчи (Locusta migratoria). Видны 11 бивалентов с одной или двумя хиазмами в каждом. На схеме (справа) отцовские и материнские хромосомы изображены соответственно сплошными и прерывистыми линиями. В каждой хиазме произошел обмен генетическим материалом. Форма бивалентов варьирует от палочковидной до крестообразной или кольцевидной, в зависимости от числа хиазм и их расположения. На этой стадии Х-хромосома интенсивно окрашивается (Д-р S. A. Henderson).
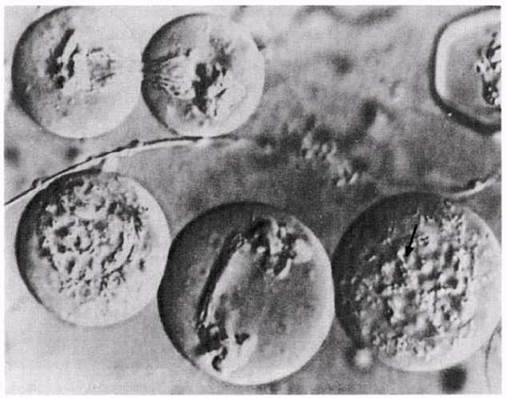
Рис. 23.14. Мейоз в живых клетках. Конъюгация и клеточное деление в живых сперматоцитах странствующей саранчи (Locusta migratoria). Препараты сфотографированы методом интерференционного контраста Номарского; этот метод с использованием поляризованного света позволяет получать удивительно объемные картины живых неокрашенных клеток. В двух клетках можно видеть конъюгацию хромосом в ранней профазе I (указано стрелкой). Две клетки (вверху слева) заканчивают первое деление мейоза. После того, как образовались две полярные группы, начинается деление всей клетки. Образуются две дочерние клетки примерно одинаковой величины. Нитевидные структуры, тянущиеся от клетки к клетке между двумя группами хромосом — это микротрубочки веретена.
23.4.1. Краткие выводы
В результате мейоза из одной родительской клетки образуются четыре дочерние клетки. Каждая дочерняя клетка содержит половину числа хромосом в родительской клетке. Обычно родительская клетка бывает диплоидной, а поэтому дочерние клетки гаплоидные.
23.4.2. Значение мейоза
1. Половое размножение. Мейоз происходит у всех организмов, размножающихся половым путем. Во время оплодотворения ядра двух гамет сливаются. Каждая гамета содержит один набор хромосом (т. е. она гаплоидна, n). В результате слияния гамет образуется зигота, содержащая два набора хромосом (т. е. диплоидная, 2n). В отсутствие мейоза слияние гамет приводило бы к удвоению числа хромосом у каждого последующего поколения, возникающего в результате полового размножения. Исключение из этого правила наблюдалось бы лишь при полиплоидии (разд. 24.9). У всех организмов с половым размножением этого не происходит благодаря существованию особого клеточного деления, при котором диплоидное число хромосом (2n) сокращается до гаплоидного (n).
23.1. На рис. 23.15 схематически представлено количество ДНК, приходящееся на одну клетку при нескольких делениях ядра.
а) Какой тип деления представлен на рис. 23.15?
б) Каким стадиям соответствуют прерывистые линии W, X и Y?
в) Какому типу клеток соответствует линия Z?
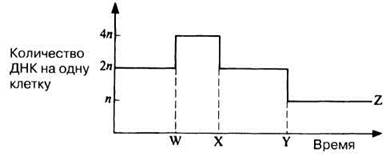
Рис. 23.15. График, который следует использовать при выполнении задания 23.1.
2. Генетическая изменчивость. Мейоз создает также возможность для возникновения в гаметах новых комбинаций генов, что ведет к генетическим изменениям в потомстве, получаемым в результате слияния гамет. В процессе мейоза это достигается двумя способами, а именно — независимым распределением хромосом и кроссинговером при первом мейотическом делении.
1) Независимое распределение хромосом. Этот процесс лучше всего объяснить на схеме (рис. 23.16). В метафазе I биваленты располагаются на экваторе веретена случайным образом. На рис. 23.16 представлена простая ситуация, в которой участвуют только два бивалента, а поэтому возможно расположение только двумя способами (при одном из них белые хромосомы лежат друг подле друга, а при другом — белая хромосома лежит подле черной). Чем больше число бивалентов, тем больше число возможных комбинаций, а, следовательно, тем выше изменчивость. Независимое распределение означает, что в анафазе I хромосомы, составляющие данный бивалент, разделяются (распределяются) независимо от хромосом других бивалентов. Черные и белые полоски на рис. 23.16 соответствуют материнским и отцовским хромосомам. Независимое распределение лежит в основе второго закона Менделя (разд. 24.1.3).
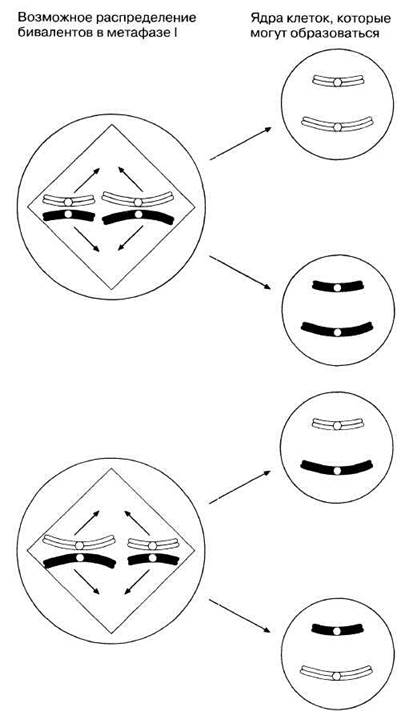
Рис. 23.16. Независимое распределение хромосом в мейозе. Показаны два бивалента, которые выстраиваются по экватору независимо от других бивалентов, так что обе возможности, представленные на схеме, одинаково вероятны. Это повышает потенциальное разнообразие гамет, которые также представлены на рисунке.
2) Кроссинговер. В результате образования хиазм между гомологичными хромосомами в профазе I происходит кроссинговер, ведущий к образованию новых комбинаций генов в хромосомах гамет. Это показано на рис. 23.11, Л, где все четыре гаметы, образовавшиеся в результате мейоза, отличаются друг от друга. Более подробно изменчивость рассматривается в разд. 24.8.4.
23.4.3. Сопоставление митоза и мейоза
Биологически значимые различия между митозом и мейозом сводятся к различиям между митозом и мейозом I. Мейоз II почти идентичен митозу. Поэтому в табл. 23.2 сравниваются только митоз и мейоз I.
Таблица 23.2. Сопоставление митоза и мейоза I
Стадия |
Митоз |
Мейоз |
Профаза |
Гомологичные хромосомы обособлены Хиазмы не образуются Кроссинговер не происходит |
Гомологичные хромосомы конъюгируют Хиазмы образуются Кроссинговер возможен |
Метафаза |
Пары хроматид располагаются на экваторе веретена |
Пары хромосом располагаются на экваторе веретена |
Анафаза |
Центромеры делятся Расходятся хроматиды Расходящиеся хроматиды идентичны |
Центромеры не делятся Расходятся целые хромосомы Расходящиеся хромосомы и их хроматиды могут оказаться неидентичными в результате кроссинговера |
Телофаза |
Число хромосом в дочерних клетках то же, что и в родительских клетках |
Число хромосом в дочерних клетках вдвое меньше, чем в родительских |
Дочерние клетки содержат обе гомологичные хромосомы (у диплоидов) |
Дочерние клетки содержат только по одной из каждой пары гомологичных хромосом |
|
Где происходит деление данного типа |
В гаплоидных, диплоидных и полиплоидных клетках При образовании соматических клеток и некоторых спор При образовании гамет у растений |
Только в диплоидных и полиплоидных клетках При образовании гамет или спор |