ГЕТЕРОФІЛІЯ У РОСЛИН - О.М. НЕДУХА - 2011
РОЗДІЛ ПЕРШИЙ. МОРФОЛОГІЧНА ТА СТРУКТУРНО-ФУНКЦІОНАЛЬНА ХАРАКТЕРИСТИКИ ВЕГЕТАТИВНИХ ОРГАНІВ ГЕТЕРОФІЛЬНИХ РОСЛИН
1.8. УЛЬТРАСТРУКТУРНІ ОЗНАКИ КЛІТИН ЛИСТКІВ І КОРЕНІВ
1.8.1. Листки Sium latifolium
S. latifolium, що зростав у воді характеризується гетерофілією: на одному пагоні розміщуються листки різної форми: підводні (двічі- або тричіпе- ристорозсічені з ниткоподібними частками), надводні (пірчасторозсічені з ланцетоподібними частками) та «перехідні» листки, у яких пластинка була над водою, а черешок - під водою (за формою пластинки подібні до підводних) (див. підрозділ 1.3.3 і рис. 1.3.1 у вставці II). Дослідження ультраструктури трьох типів листків веху широколистого показало наявність як спільних ознак, так і певні відмінності в ультраструктурі клітин мезофілу (Недуха, 2004).
Підводні листки. Аналіз ультраструктури клітин мезофілу підводних листків показав, що клітини палісади й губчастого мезофілу мали велику центральну вакуоль; цитоплазма з органелами розміщувалась уздовж клітинних оболонок. Хлоропласти палісадного й губчастого мезофілу лінзоподібної або видовженої форми (залежно від площини зрізу) мали подібну ультраструктуру. Число хлоропластів на зрізі клітини палісадної паренхіми становило 9,0 ± 2,4; на зрізі губчастої паренхіми 6,7 ± 0,3 (табл. 1.8.1.1). Середній розмір хлоропластів становив 4,3 х 1,2 мкм, мембранна система гранального типу (рис. 1.8.1.1, а, б), добре розвинута, зерна крохмалю виявлялися рідко. Характерною особливістю пластид була наявність великої кількості тилакоїдів у грані (табл. 1.8.1.2), діаметр яких коливався від 0,2 до 0,4 мкм, кількість гран на зрізі хлоропласта варіювала від 6-8 до 12-14. В деяких хлоропластів тилакоїди гран і строми характеризувалися підвищеною електронною щільністю. Зрідка в щільній стромі спостерігали пластоглобули діаметром до 50 нм. Мітохондрії мали розвинуту систему крист (рис. 1.8.1.1, в). В цитоплазмі виявлено багато рибосом, ендоплазматичний ретикулум гранулярного типу. Ширина клітинних оболонок становила 0,2-0,4 мкм. Ядро округле чи злегка витягнуте, нукле- оплазма щільна, в ядерці виявляли гранулярний та фібрилярний компоненти (рис. 1.8.1.1, г).
«Перехідні» листки. Клітини мезофілу перехідних листків (черешок яких був занурений у воду, а пластинка - над водою) відрізнялись за ультраструктурними ознаками. Умовно ми поділили їх на два типи: в одних ультраструктура ендомембран і органел характеризувалася інтактністю (рис. 1.8.1.2, а, б), в інших - спостерігали деструкцію органел і ендомемб- ран (рис. 1.8.1.2, в, г).
В клітинах першого типу мітохондрії були конденсованого типу, апарат Гольджі представлений групами коротких диктіосом, ендоплазматичний ретикулум гранулярний. В гіалоплазмі видно рибосоми й полісоми.
Ширина клітинної оболонки становила 0,3-0,4 мкм. Хлоропласти відрізнялися від таких в клітинах попереднього варіанта лінійними розмірами, кількістю зерен крохмалю (табл. 1.8.1.1) і тилакоїдів в гранах (табл. 1.8.1.2); гран з 20 і більшою кількістю тилакоїдів було майже в 2,5 раза менше, ніж в хлоропластах підводних листків.
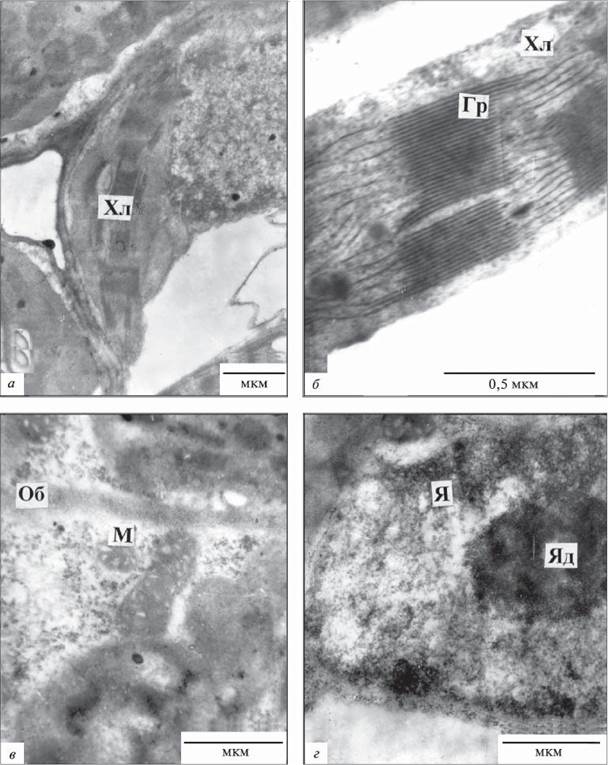
Рис.1.8.1.1. Фрагменти клітин мезофілу підводних листків Sium latifolium:
М - мітохондрія, Хл - хлоропласт, Гр - грана, Об - клітинна оболонка, Я - ядро, Яд - ядерце;
а,б - палісадна паренхіма, в, г - губчаста паренхіма
Особливістю таких клітин була наявність майже округлих електронно- щільних утворень розміром 10-15 нм, що контактували з мембранами оболонок органел і цитоплазматичною мембраною (рис. 1.8.1.2, а, б). В клітинах другого типу (рис. 1.8.1.2, в, г) виявлено чіткі ознаки деструкції, характерні для клітин, що старіють: порушення інактності цитоплазматичної мембрани й тонопласту, деструкція нуклеоплазми й ущільнення ядерця, в цитоплазмі - підвищена електронна щільність органел.
ТАБЛИЦЯ 1.8.1.1. Кількісна характеристика хлоропластів клітин мезофілу Sium latifolium
Тип листка |
Кількість хлоропластів на зрізі різних клітин мезофілу |
Розмір хлоропласта, мкм |
Розмір крохмального зерна, мкм |
Кількість зерен крохмалю на зрізі хлоропласта |
|||
Палісадний |
губчастий |
довга- вісь |
коротка- вісь |
довга- вісь |
коротка- вісь |
||
Підводний |
9,0 ± 2,4 |
6,7 ± 0,3 |
4,3 ± 0,3 |
1,2 ± 0,1 |
1,4 ± 0,1 |
0,6 ± 0,19 |
0,7 ± 0,1 |
«Перехідний» |
10 ± 0,1 |
6,5 ± 0,3 |
6,0 ± 0,2 |
1,9 ± 0,1 |
1,3 ± 0,1 |
0,5 ± 0,1 |
2,3 ± 0,2 |
Надводний |
7,5 ± 0,3 |
6,6 ± 0,4 |
4,4 ± 0,1 |
1,3 ± 0,1 |
1,2 ± 0,1 |
0,3 ± 0,02 |
1,1 ± 0,1 |
ТАБЛИЦЯ 1.8.1.2. Кількісна характеристика тилакоїдів хлоропластів мезофілу Sium latifolium
Тип листка |
Розподіл гран за кількістю тилакоїдів, % |
||||
2 - 3 |
4 - 6 |
7 - 10 |
10 - 20 |
понад 20 |
|
Підводний |
- |
8,6 |
26,6 |
30,0 |
34,8 |
«Перехідний» |
11,1 |
18,5 |
14,8 |
40,8 |
14,7 |
Надводний |
5,3 |
66,2 |
28,5 |
- |
- |
ТАБЛИЦЯ 1.8.1.3. Вміст пігментів у листкових пластинках Sium latifolium
Пігменти |
Листок |
||
підводний |
«перехідний » |
надводний |
|
Хлорофіл а |
|||
мг/г сирої маси |
0,654 |
0,411 |
1,053 |
мг/г сухої маси |
1,712 |
2,020 |
1,316 |
Хлорофіл b |
|||
мг/г сирої маси |
0,443 |
0,315 |
0,391 |
мг/г сухої маси |
1,180 |
1,298 |
0,510 |
Сума хлорофілів (a+b) |
|||
мг/г сирої маси |
1,097 |
0,726 |
0,391 |
мг/г сухої маси |
2,892 |
3,318 |
1,826 |
Каротиноїди |
|||
мг/г сирої маси |
1,179 |
0,114 |
0,598 |
мг/г сухої маси |
3,148 |
0,310 |
0,780 |
Відношення хлорофілу а/b |
|||
на сиру масу |
1,48 |
1,31 |
2,69 |
на суху масу |
1,45 |
1,55 |
2,58 |
Хлоропласти характеризувались щільною стромою або ж її повною просвітленістю, в стромі спостерігали зерна крохмалю й пластоглобули; зруйновані тилакоїди гран і строми мали високу електронну щільність. В стромі хлоропластів було багато пластоглобул, рибосом не спостерігали. Гранулярний ендоплазматичний ретикулум, рибосоми й полісоми не виявлялися. В таких клітинах спостерігали округлі або овальні електронно- щільні утворення (розміром 10-100 нм), що контактували з ендомембра- нами: оболонками органел, цитоплазматичною мембраною й ендоплазматичним ретикулумом (рис. 1.8.1.2, в, г). Такі формування за локалізацією та розмірами нагадують кальцієві сайти в клітинах коренів і листків Alisma plantago-aquatica, коренів гороху (Белявская, 1994; 2003; Кордюм и др., 2003).
Надводні листки. Органели в клітинах мезофілу S. latifolium розміщувались уздовж оболонок (рис. 1.8.1.3, а-г). Форма хлоропластів в клітинах палісадної й губчастої паренхіми залежно від площини зрізу була лінзоподібною або злегка видовженою (рис. 1.8.1.3, б, г). Хлоропласти містили крохмальні зерна (табл.1.8.1.1); численні грани складалися з невеликої кількості тилакоїдів (табл. 1.8.1.2) з діаметром 0,2-0,5 мкм. В щільній стромі пластид видно рибосоми.

Рис. 1.8.1.2. Фрагменти клітин мезофілу «перехідних» листків Sium latifolium: М - мітохондрія, Хл - хлоропласт, Об - клітинна оболонка, Я - ядро; а, б - губчаста паренхіма, в, г - палісадна паренхіма
Мітохондрії конденсованого типу, округлі або злегка витягнуті, їх розмір по довгій осі становив 0,5-0,8 мкм. Часто спостерігали контакти мі- тохондрій з хлоропластами. В цитоплазмі багато рибосом; короткі канали гранулярного ендоплазматичного ретикулуму розміщувались уздовж оболонок. Апарат Гольджі був представлений групами диктіосом з 3-4 цистерн і кількох пухирців. Ядро округле чи овальне з нещільною нукле- оплазмою (рис. 1.8.1.3, а, г).
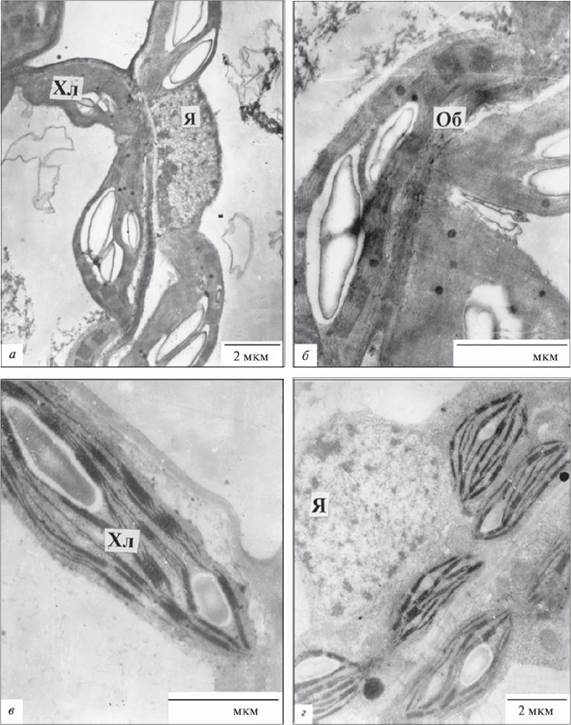
Рис. 1.8.1.3. Фрагменти клітин мезофілу надводних листків Sium latifolium: Хл - хлоропласт, Об - клітинна оболонка, Я - ядро; а, в - палісадна паренхіма, б, г - губчаста паренхіма
Вміст фотосинтезуючих пігментів, в т.ч. хлорофілів а та b, а також каротиноїдів, відрізнявся в підводних і надводних листках (табл. 1.8.1.3). Якщо порівнювати їх вміст на сиру масу, то вміст хлорофілу а в надводних листках був майже удвічі більшим порівняно з таким в підводних, тимчасом як його вміст на суху масу був більшим в підводних і «перехідних» листках. Вміст хлорофілу b (на суху масу) був вищим в підводних і перехідних листках. Визначено, що сума хлорофілів а + b (на суху та сиру масу) також була достовірно більшою в підводних і «перехідних» листках порівняно з надводними. Відношення хлорофілу а до хлорофілу b було в 1,7-1,9 раза більшим (на сиру й суху масу) в надводних листках.
Отже, ультраструктурний аналіз клітин листків гетерофільної рослини - веху широколистого показав, що структура клітин мезофілу підводних і надводних листочків веху типова для клітин мезофітних суходільних рослин (Гамалей, Куликов, 1978). Однак ми виявили специфічні риси структури, характерні лише для підводних листків. Перша особливість - це наявність гран з великою кількістю тилакоїдів. Як відомо, ця ознака є типовою для хлоропластів листкових пластинок тіньовитривалих рослин і зумовлена посиленим синтезом хлорофілу b (Горьішина, 1989; Николаева, Власова, 1990). З урахуванням того, що водна поверхня частково відбиває та поглинає світло (Ландсберг, 1986), можна констатувати, що підводні листки водного екотипу веху одержували значно менше світла, ніж надводні, що впливало на їхню структуру.
Другою особливістю структури хлоропластів підводних листків веху широколистого, що відрізняє їх від хлоропластів надводних листків, є менша кількість зерен крохмалю в пластиді. Кількість зерен крохмалю в хлоропластах підводних листків була в 1,7 раза нижчою, ніж в хлоропластах надводних листків і утричі меншою порівняно з хлоропластами «перехідних» листків. Ми припускаємо, що для підводних листків характерне зниження синтезу крохмалю. Це підтверджується даними Г.Ф. Некрасової з співав., які радіоізотопним методом вивчали інтенсивність фіксації СО2 та вміст продуктів фотосинтезу в підводних і надводних листках 42 видів гідрофітів і встановили, що в рослинах із зануреними у воду листками було суттєво меншим включення 14С в цукрозу й крохмаль (в середньому, відповідно, 30 і 9%) порівняно з надводними й плаваючими (45 і 15%) листками, водночас у них виявлено в 1,5 раза більше 14С у С4-кислотах (Некрасова, и др., 2003).
Ще одна виявлена нами особливість підводних листків веху - високий вміст хлорофілу b і знижене майже удвічі відношення хлорофілу а/ b порівняно з такими же показниками надводних листків. Як відомо, ці ознаки характерні для рослин, що ростуть у затінку та в умовах низької освітленості (Силаева, 1978; Горьішина, 1989) З урахуванням цих даних і відомостей літератури про те, що хлорофіл b переважно зосереджується у тилакоїдах гран (Гудин, Мерсер, 1986), можна говорити про кореляцію структури хлоропластів підводних листків веху й вмісту в них пігментів.
Описані нами часткова деструкція мембран (тонопласту й плазмалеми), зниження чи повна відсутність рибосом в цитоплазмі певної частини клітин мезофілу «перехідних» листків є типовими для листків, що старіють (Гамалей, Куликов, 1978), та для мезофілу листків в умовах впливу на рослину несприятливих факторів (Силаева, 1978). Виявлені нами в таких клітинах електроннощільні утворення за формою, розмірами та локалізацією подібні глобулярним утворенням, відомим у літературі як кальцієві сайти рослинних та тваринних клітин (Белявская, 1994; 2003). Останні утворюються в разі підвищення рівня цитоплазматичного кальцію вище фізіологічного як внаслідок інгібування активності Са2+-АТФази плазма- леми, так і при активації перекисного окислення ліпідів цитоплазматичної мембрани, що призводить до порушення кальцієвого балансу клітин та їх старіння (Roux, Slocum, 1982; Белявская, 1994; 2003) .
Поява великої кількості добре розвинутих мітохондрій в клітинах мезофілу підводних і перехідних (плаваючих) листків в рослин веху на фазі вегетативного розвитку свідчить про те, що інтенсивне дихання відбувається одночасно з фотосинтезом, забезпечуючи клітини утворенням АТФ, що необхідно для синтезу цукрів і полісахаридів при рості рослини (Kromer, 1995). Крім того, збільшення популяції мітохондрій в клітинах мезофілу перехідних листків з ознаками деструкції інших ор- ганел, очевидно, може свідчити про індукцію запрограмованої смерті у відповідь на збільшення в клітинах активних форм кисню. Подібний феномен описаний раніше в клітинах Arabidopsis thaliana (L.) Heynh., що призводило до виснаження АТФ, окисного вибуху й наступної клітинної смерті (Keiko Yoshinaga et al., 2002; Maike et al., 2004). Поява в клітинах мезофілу перехідних листків веху широколистого великої кількості пероксжом, що контактують з хлоропластами, очевидно, свідчить про посилення фотодихання в клітинах перехідних листків поряд з ознаками старіння.