ГЕТЕРОФІЛІЯ У РОСЛИН - О.М. НЕДУХА - 2011
1.3. ФОРМА ЛИСТКІВ ГЕТЕРОФІЛЬНИХ РОСЛИН
У вищих водних гетерофільних рослин підводні та надводні листки мають різну форму. Форму листків ми описували згідно морфологічного опису листків за А.Л. Федоровим та співав. (1956). Умовно ми поділили підводні листки на три типи: розсічені, лінійні (видовжені) і округлі. Розсічений (непарнопірчасті) тип листкових пластинок трапляється в Sium latifolium, Proserpinaca palustris (Schmidt, Millington, 1968), Myriophyllum sp., Trapa natans, Potamogeton sp. (Frost-Christensen, Sand-Jensen, 1995; Arber, 2008). Підводні лінійні листки характерні для всіх видів стрілолиста (Sagittaria sp.) (Arber, 2008), овальні чи округлі придонні з дуже коротким черешком - у видів родини Nymphaeaceae (Лататтєві): глечики Nuphar lutea, латаття сніжнобіле Nymphaea candida, N. adorata Aiton та ін. (Kane, 1984; Villani, Etnier, 2008). Розсічена та сильно видовжена форма підводних листків допомагає протистояти опору та тиску води як у стоячій воді, так і в річках при течії та дії хвиль (Tsukaya, 2006).
Одним із прикладів підводних розсічених листків є листки багаторічника Sium latifolium (Umbrelliferaceae), у якого навесні (у травні) листки за формою та розміщенням на стеблі дуже різняться (рис. 1.3.1, див. вставку ІІ). Підводні листки повністю занурені у воду, двічі- (або тричіпірчасторозсічені з ниткоподібними частками (рис. 1.3.1, а). Листкові пластинки підводних листків складалися з 6-8 парних часток та однієї непарної; середній розмір довгої осі листкової пластинки - 10-12, короткої - 7-9 см; середній розмір ширини листкових часток коливався від 0,8 до 3 мм. Довжина черешка залежала від глибини зростання, і коливалась від 8 до 13 см.
Листки другого порядку S. latifolium за формою подібні до підводних; черешки цих листків занурені у воду, а пластинки - над водою. Листки складаються із 7-8 парних та однієї непарної листкових часток, які удвічі ширші за частки листків першого порядку (рис. 1.3.1, б). Довжина черешка в листків другого порядку коливалась від 10 до 20 см.
Листки третього й четвертого порядку S. latifolium - надводні, середній розмір пластинок більший, ніж у підводних листків. Довга вісь листка коливалась від 16 до 18 см, коротка - 10-14 см; довжина черешків коливалась від 15 до 19 см. Надводні листки пірчасторозсічені, їхні частки 4-6-парні, овальні (нерівнобокі біля основи) дрібногостропильчасті (Недуха, 2010, а). Розмір довгої осі листкових часток змінювався від 5 до 7 см, короткої - від 2 до 2,6 см. Sium latifolium - багаторічники з повзучими підземними пагонами. Зубці чашечки малопомітні. Пелюстки білі.
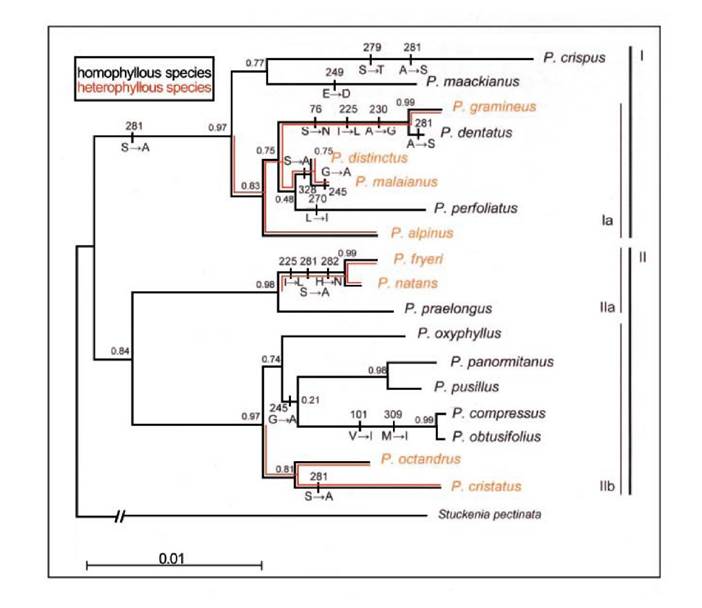
Рис. 1.2.4. Філогенетичне cpDNA дерево роду Potamogeton, реконструйоване на базі даних комбінації 4394-bр хлоропластної ДНК. Виміри вказують на підтримку галужень (Anisimova, Gascuel, 2006). Заміни амінокислотних залишків у гені RbcL реконструйовано на моделі підстановки кодону (М8). Галуження, які вказані оранжевим кольором - це три гетерофільні лінії; галуження, що вказані чорним кольором - це гомофільні лінії видів роду Potamogeton (Iida et al., 2009)

Рис. 1.3.1. Загальний вигляд листків гетерофільних повітряно-водних рослин Sium latifolium (a-в). Підводні листки тричіпірчасторозсічені (б). Листки другого порядку, черешок яких був у воді, а пластинка - над водою також мали двічіпірчасторозсічену форму (в), їх частки більшого розміру, ніж частки підводних листків. Надводні листки (а - б) непарнопірчастсторозсічені, їхні частки мали овальну форму. Матеріал зібрано у травні 2007 р. (Конча-Заспа, під Києвом)

Рис. 1.3.2. Trapa natans характеризується двома формами підводних листків [(лінійні (а) і розсічені (б-г)] і рівномірно трикутними плаваючими листками (б і в). При проростанні плодів на підводному пагоні спочатку утворюються 4-5 лінійних листка (а). Коли пагін із розеткою надводних листків виходить на водну поверхню, на підводному пагоні формуються розсічені підводні листки (б, в), які з часом сильно збільшуються в розмірі (г). Черешки плаваючих листків мають розширення ("здуття"), що містить повітряні порожнини. Підводні лінійні і розсічені (а - г) листки зелені, поряд корені - рожеві (г). Матеріал зібрано на Русанівському каналі, лівий берег Дніпра (район Києва) у травні-червні 2007-2011 р.

Рис. 1.3.3. Загальний вигляд Sagittaria sagitHfolia (а, б) і S. filiformis (г). Підводні листки лінійної форми (б - г); надводні - стрілоподібної (а, б, в), продовгуватої (б, в) і ланцетоподібної (г). Стрілолист звичайний (рис. б, в) зібрано на берегах р. Псьол біля с.м.т. Велика Багачка Полтавської області в червні 2010 р.; (рис. г - стрілолист ниткоподібний, http://ru.wikipedia.org/wiki/)

Рис. 1.3.4. Загальний вигляд Nuphar lutea. Плаваючі листки (a-в) мали серцевидноовальну форму, підводні листки - округлу форму (а, б). Матеріал зібрано в червні 2008 р. на р. Псьол біля с.м.т. Велика Багачка Полтавської обл. (рис. а і б) і на Русанівському каналі р. Дніпро (Київ) (рис. в)

Рис. 1.3.5. АБК-залежна гетерофілія у рослини Marsilea quadrifolia спричинена дією АБК (1 мМ): а - загальний вигляд рослини, б - підводний листок, в - надводний листок (Bai-Ling Lin, 2002)

Рис. 1.3.6. Загальний вигляд листків суходільної екоформи Sium latifolium - веху широколистого на стадії вегетативного росту. Перші три листки мали цілісну пластинку (а, б), наступні - пірчасторозсічену (в). Матеріал зібрано у травні 2007 р. (Конча-Заспа під Києвом)

Рис. 1.3.8. Послідовний ряд листків прикореневої розетки Pastinaca sativa: l-з першим справжнім листком; 2 - відмираючий листок з трьома лопатями; З - трійчатоскладний листок з відмираючими листочками; 4-6 - трійчатоскладні листки різних ярусів; 7-8 - непарноперистоскладні листки (Наумов, 2009)

Рис. 1.3.9. Загальний вигляд Campanula rotundifolia: а - прикореневі листки загостренояйцеподібні; б, в - стеблові листки лінійні. (Фото взято: USDA, US Dep. of Agricultur Natural Res. Conservation Service, www.rook. org/.../ companularot.html)

Рис. 1.3.10. Загальний вигляд трьох типів листків Triphyophyllum peltatum: а - прикореневі списоподібні листки; б - стеблові лінійні, комахоїдні листки із залозками; в - стеблові листки лопатчаті, з двома крючкоподібними виростами на кінчику пластинки. (Фото взято: http://en.wikipedia. org/wiki/triphyophyllumi http://www.righthealth.com/topic/triphyophyllum)

Рис. 1.6.1.1. Загальний вигляд листків Sium latifolium: 1- підводний листок, 2 - «перехідний" листок, 3 - надводний листок. Рослини збирали в травні, (Конча-Заспа під Києвом)
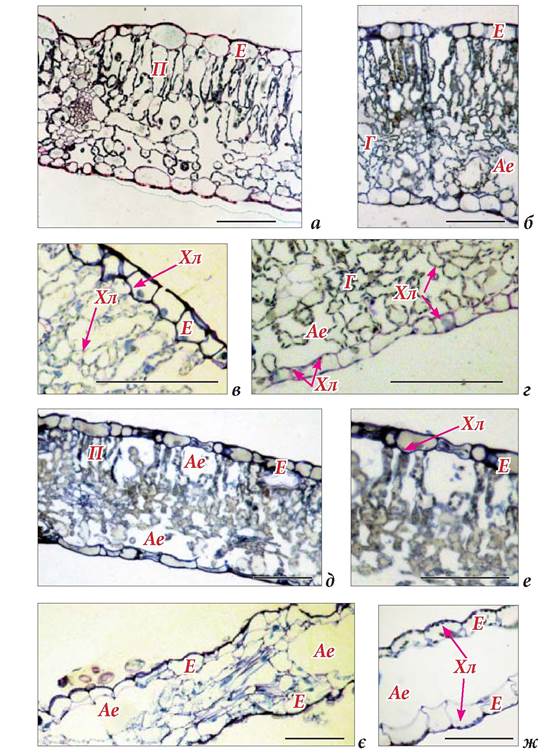
Рис. 1.6.2.1. Поперечні зрізи листкових пластинок Sagittaria sagittifolia: а-г - надводний стрілоподібний листок, д, Є - надводний продовгуватий листок; Є, Ж- підводний лінійний листок. Стрілками позначені хлоропласти. Матеріал зібрано на березі р. Псьол (с.м.т. Велика Багачка Полтавської області). Позначення: Ае - аеренхіма, Г - губчаста паренхіма, Е - епідерміс, П - палісадна паренхіма, Хл - хлоропласт. Масштаб = 100 мкм

Рис. 1.6/3.1. Поперечні зрізи плаваючого (а, г, д) і підводного (б, в) листків Nuphar lutea, фаза бутонізації. На рисунках г і д збільшені фрагменти плаваючого листка, що обведені квадратами на рисунку а. На рис. в збільшений фрагмент підводного листка, що обведено квадратом на рисунку б. Листки глечиків зібрані на р. Псьол (с.м.т. Велика Багачка Полтавської області). Позначення: Ае - аеренхіма, Г - губчаста паренхіма, Е - епідерміс, П - палісадна паренхіма, ФП - фотосинтезуюча паренхіма підводних листків. Масштаб = 100 мкм
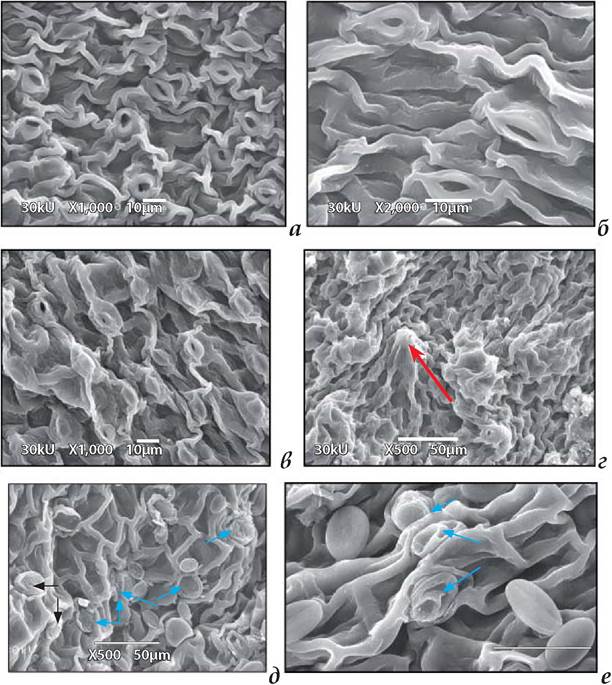
Рис. 1.7.5.3.1. Структура верхньої (а, б) та нижньої (в-е) поверхонь плаваючих листків Nuphar lutea. В епідермі видно гідропоти (вказано стрілками на рис. д, е) і одноклітинні водорості, що прикріплені до кутикули епідерми. Масштаб: а,б,в = 10 мкм, г, д, е = 50 мкм
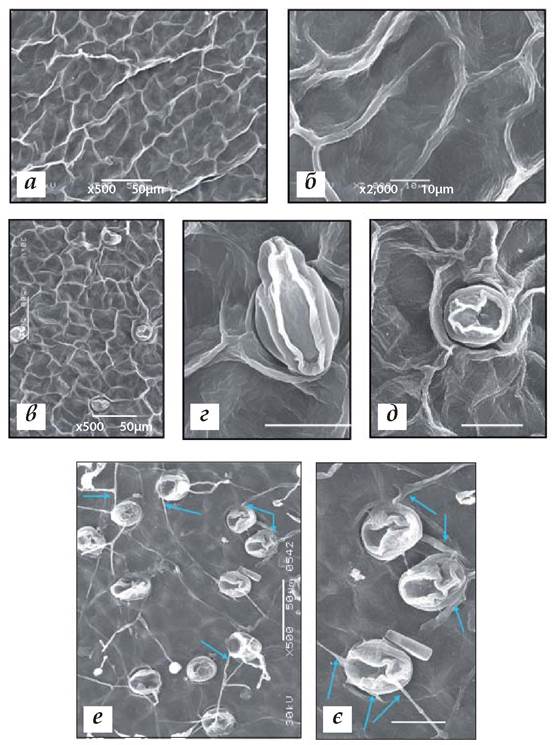
Рис. 1.7.5.3.2. Структура верхньої (а, б) і нижньої (в-є) поверхонь підводних (придонних) листків Nuphar lutea. На нижній поверхні видно гідропоти (рис. в-є) чашоподібної форми, що з'єднані між собою й основними клітинами епідерми щупальцеподібними кутикулярними тяжами (рис. е, є, стрілки). Підводні листки рослин зростали на глибині біля 2 метрів від водної поверхні (на р. Псьол). Червень, 2008 р., р. Псьол біля с.м.т. Велика Багачка Полтавської обл. Масштаб: а, в, е = 50 мкм; 6 = 10 мкм; г, д, є = 20 мкм

Рис. 2.2.2.1. Загальний вигляд листків повітряно-водних (а) та суходільних рослин (б) Sium latifolium. Різна форма пластинок у повітряно-водних рослин: у підводних і перехідних листків (тричіпірчасторозсічена); у надводних листків - пірчасторозсічена (рис. а). Листки суходільних рослин мають дві форми пластинок (б) : перші три листка - цілісну загострено яйцеподібну пластинку, листки наступних ярусів — пірчасторозсічену
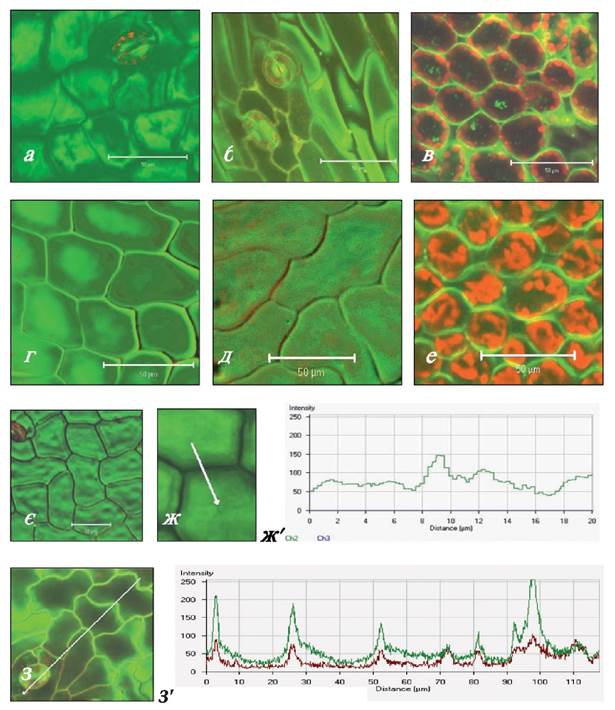
Рис. 2.2.2.2. Клітини епідермісу (а, б, г, д, є-з) та мезофілу (в, е) листків повітряно-водних (а-д, ж, з) та суходільних (е, є) рослин Sium latifolium після інкубації в розчині калькофлуору (кф.) Комплекс целюлоза+кф. флуоресціює зеленим кольором, автофлуоресценція хлорофілу червоного кольору, ж', з' - діаграми інтенсивності флуоресценції комплексу целюлоза + кф. (зелена крива) та автофлуоресценції хлорофілу (червона крива). По горизонталі - відстань (мкм), що була просканована, позначена білою стрілкою (ж, з), по вертикалі - інтенсивність флуоресценції в умовних одиницях.
Масштаб =50 мкм
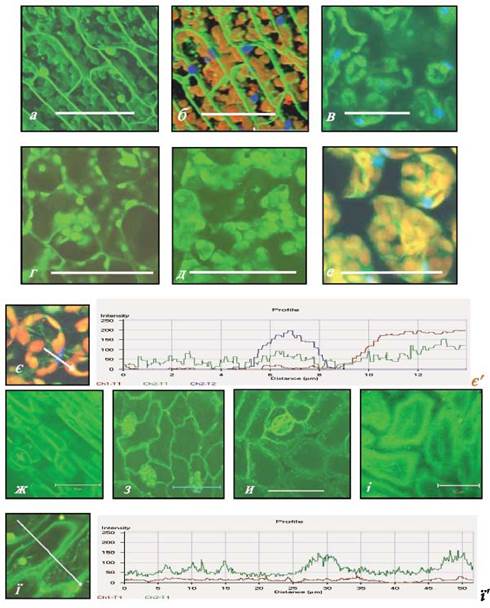
Рис. 2.2.3.1. Клітини мезофілу (а-е) та епідермісу (ж-і) листків Sium latifolium після інкубації в розчині Fluo-4 (комплекс Са2+ + Fluo-4 флуоресціює зеленим кольором) і після інкубації в розчині DAPI (ДНК ядер флуоресціює блакитним кольором), автофлуоресценція хлорофілу має червоний колір. Листки повітряно-водних рослин виду: а, б, в, є, ж, з - підводні; г, и - надводні. Листки суходільних рослин S. latifolium: д - з цілісною пластинкою; е, і, ї - з розсіченою пластинкою, а, б - клітини палісадної паренхіми, в-е - клітини губчастої паренхіми; є', ї' - діаграми інтенсивності флюоресценції комплексу Са2+ + Fluo-4 (зелена лінія), автофлуоресценції хлорофілу (червона лінія), флуоресценції ДНК (блакитна лінія) в клітинах мезофілу (є') підводних листків повітряно-водної рослини та адаксіальної поверхні епідермісу (V) листків суходільної рослини. По горизонталі: відстань (мкм), що була просканована, позначена білою стрілкою на рисунках є і ї; по вертикалі: інтенсивність флуоресценції комплексу Са2+ + Fluo-4 в умовних одиницях. Масштаб = 50 мкм
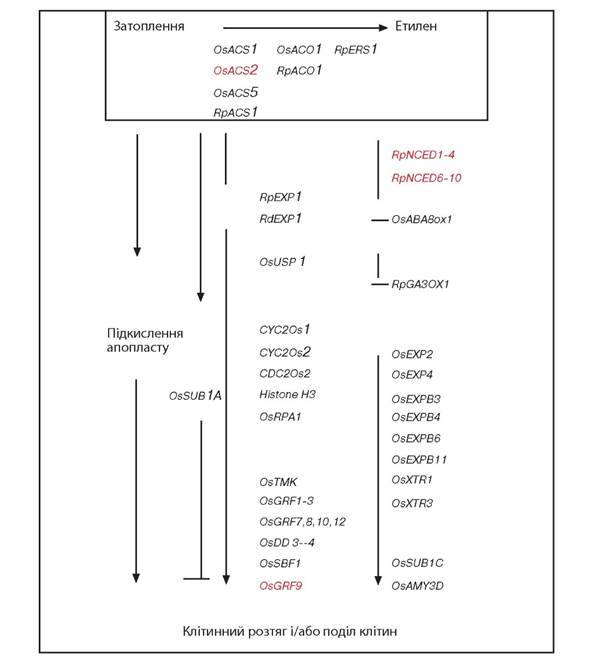
Рис. 2.2.4.1. Схематична зображення експресії генів, залучених до поділу й розтягування клітин стебел при затопленні рослин (чорним кольором вказано на стимулюючу дію генів, червоним - на інгібуючу) (Bailey-Serres, Voessenek, 2008)
Абревіатура генів, які індукують синтез наступних білків :
CyC2Os - циклін; CDC2Os - циклін-залежна кіназа; OsACO та RpACO - ACC-оксидаза; OsACS та RpACO - ACC-синтетаза; OsDD - білок диференціації (Kende et al., 1998); OsAMY - амілаза (Fukao et al., 2006); OsEXP, RdEXP та RpEXP - експансини; OsGRF - рост-регулюючий фактор (Choi et al., 2004); OsRPA - реплікуючий білок А1; OsSBF - білок із родини симпорту натрій-жовчна кислота (Rzewuski, Sauter, 2002); OsSUBl - білок підводний; OsTMK - трансмембранна протеін кіназа (Van der Knaap et al., 1999); OsUSP - універсальний стресовий білок (Sauter et al., 2002); RpERS1 - рецептор етилену (Vriezen et al, 1997); RpNCED - 9-цисепоксикаротиноїд діоксигеназа; RpGA3ax - гіберелін 3-оксидаза (Benschop et al., 2006); OsXTR - ксилоглюкан ендотрансглюкозилаза зв'язана (Darley et al., 2001); OsABA8ox - АБК 8'-гідролаза (Saika et al., 2007). Позначення: Os - Oryza sativa; Rd - Regnellidium diphyllum; та Rp - Rumex palustris або Ranunculus palustris. OsACS1 - синтетаза, OsACO1 - оксидаза в Oryza sativa, RpERS1 - етилен-рецептор-синтетаза (Vriezen et al., 1997) в Ranunculus palustris
Плід яйцеподібний або довгастий, стиснутий із боків, із ниткоподібними ребрами, ребра плоду вузькі. Стовпчик вільний.
Підводні листки Trapa natas L. мають різну форму: лінійну і розсічену; плаваючі листки - рівносторонню трикутну (рис. 1.3.2, див. вставку ІІІ). Першими на проростаючому підводному пагоні (у квітні-травні) з'являються 4-5 сидячих лінійних листка (20-30 мм висотою і біля 2-2,5 мм шириною кожний) з гострою верхівкою; краї цільні. Коли пагін (кінець травня-червень) досягне водної поверхні, починає роспускатися розетка плаваючих листків, кількість яких в розетці варіює від 9 до 18. На цей час перший тип підводних лінійних листків починає відмирати, і на нижній частині пагона (на віддалі 10-20 мм від плода) починає формуватися другий тип підводних листків - три-пять розсічених листків з коротким черешком; довжина листків 4-6,5 см, ширина 2-4,5 см; листок містить 15-47 сегментів. Плаваючі листки мають рівносторонню трикутну форму, основа листка - округла-клиновидна (рис. 1.3.2, див. вставку ІІІ), вершина - гостра, краї листків - зубчасті; черешок циліндричний (довжиною 2-4 см, діаметром 2-2,5 мм), посередині має «здуття», розмір якого у 1,5-1,9 раза перевищує діаметр черешка біля основи. Здута частина черешка містить розвинену аеренхіму, завдяки якій листок плаває на водній поверхні. T. natas - це однорічна водна рослина з пучками нитчатоподібних коренів світло-рожевого (квітень-травень) або світло- коричневого кольору (травень-вересень), тонким галузистим стеблом, довжина якого варіює (залежно від глибини водоймища) від 40 см до 200 і більше см (рис. 1.3.2, б). Від стебла галузяться корені (рис. 1.3.2, б, г). В середині червня плід (горіх) відділяється від рослини.
Плаваючі листки Trapa natans, біля основи мають лінійні або вузько ланцетні прилистки. Квітки дрібні, двостатеві, правильні, з подвійною оцвітиною розміщені поодинці в пазухах листків. Пелюсток чотири, білого кольору. Плід - кістянка, з тонким оплоднем і дерев'янистою оболонкою, що має 2-4 роговидні вирости. Крохмалисте насіння водяного горіха містить близько 15% білків, 7,5% жиру, 52% крохмалю та 3% цукру, насіння їстівне в сирому та вареному вигляді, розмелюється на крупу чи борошно.
Лінійні, видовжені або шаблеподібні підводні листки характерні для більшості видів родини Alismataceae. Так, у Sagittaria sagittifolia підводні листки лінійної форми; листкова пластинка повітряних листків має стрілоподібну і продовгувату форму (рис. 1.3.3, див. вставку ІУ). S. sagittifolia - багаторічна рослина з білими квітками, видовженим кореневищем, від якого відходять численні корені. S. sagittifolia росте на різній глибині - від кількох десятків см до 1,5 м на мулистому грунті.
Підводні листки стрілолиста звичайного сидячі, завдовжки 50-80 см, на мілководді значно коротші. Ширина підводної листкової пластинки варіює від 2 до 4 см і більше, довжина - також варіює залежно від глибини зростання стрілолиста, жилкування підводних листків дугоподібне. Підводні листки світло-зелені, з'являються після проростання насіння або утворюються із бруньок на підземному пагоні. Навесні вони швидко ростуть, але не досягають поверхні води. Надводні листки темно-зеленого кольору разом із черешками досягають 50 см чи/і більше. Сама пластинка має стрілоподібну форму: з гострою верхівкою та гострими базальними лопатями, які розходяться.
У стрілолиста звичайного, який збирали в Полтавській області вздовж берегів р. Псьол, у надводних листків ми виявили дві форми листкових пластинок, стрілоподібну та продовгувату (рис. 1.3.3, а-в, див. вставку ІУ). У рослин цього виду було 2-7 надводних листків зі стрілоподібною пластинкою та один-два листки - з продовгуватою пластинкою (рис. 1.3.3, б, в), а також 5-12 підводних видовжених листків лінійної форми. Квітки розвиваються над поверхнею води, з білою оцвітиною, біля основи темно-фіолетові чи рожеві, трипелюсткові, у мутовці по дві чи три квітки, у нижній частині суцвіття розміщені жіночі квітки з короткими квітконіжками, а зверху - чоловічі з довгими квітконіжками. Тичинок шість. Для надводних листків Sagittaria filiformis (рис. 1.3.3, г, див. вставку ІУ) характерна ланцетоподібна форма, для підводних листків - лінійна форма, аналогічно формі підводних листків у S. sagittifolia. У стрілолиста збірний плід з шести-восьми, зірчасто розміщених, не сплюснутих листянок. Квітколоже при плодах видовжене. Плодики 6-10 мм завдовжки.
Третій тип форми підводних листків, який ми виділили у рослин, які характеризуються гетерофілією - це придонні округлі листки, які трапляються у видів родини Nymphaeaceae. Наприклад, у Nuphar lutea підводні листки округлі, по краю складчасті, дуже тонкі, майже «прозорі». Надводні листки серцевидноовальної форми (рис. 1.3.4, див. вставку У). або яйцевидноовальної, товсті, без прилистків, з тригранними черешками, довжина яких залежить від глибини зростання рослини, у підводних листків довжина черешка становить 7-15 см. Чашечка п'ятилиста, пелюстки численні, дрібніші за чашолистки, жовті. Квітки 4-5 см діаметром. На Україні квітне з червня по вересень. Поверхня плоду гладенька (Визначник рослин УРСР, 1950).
Крім зазначених трьох типів форми підводних листків, у природі трапляються також й інші форми (рис. 1.3.5, див. вставку VI).). Надводні листкові пластинки в Marsilea quadrifolia мають колоподібну форму, діаметром близько 10 мм, пластинка розсічена (до половини радіуса) навхрест під кутом 90о на чотири лопаті, краї лопатей пластинки округлі (рис. 1.3.5, а, в). Підводні листкові пластинки в M. quadrifolia також чотирьохрозсічені, складаються з чотирьох сегментів (рис. 1.3.5, а, б) розміром кілька мм, які відходять від короткого черешка (0,7-1,5 см), краї сегментів округлі (Ваі-Ling Lin, 2002).
Гетерофілія в суходільних рослин залежить від виду й проявляється в онтогенезі: під час росту вегетативного пагона, особливо на ранніх стадіях його розвитку, а також при формуванні квітконосного пагона, або ж коли у багаторічних рослин листки формуються у різні сезони (Сацьіперова, 1984; Наумов, 2009; Givnish et al., 1994; Eckenwalder, 1980; Webb, 1984; Christodoulakis, 1989; Christodoulakis et al., 1990). За ранніми даними К. Ге- бела (Goebel, 1891; 1900), який для опису розвитку рослин запропонував термін гетеробластний розвиток, різна форма листків у суходільної рослини Hedera helix проявлялася весною: перші ювенільні листки мали перисту форму пластинки, а наступні листки - цілісну листкову пластинку (Goebel, 1990; цит.: за Schmidt, Millington, 1968).
Умовно ми розділили суходільні гетерофільні види рослин за формою листкових пластинок на два типи: перший тип - види, у яких форма листків ускладнюється (від цілісної пластинки до розсіченої чи ін.) при формуванні вегетативного пагона, або ж навпаки, форма спрощується при формуванні квітконосного пагона; другий тип - види, у яких листки прикореневої розетки за формою відрізняються від стеблових листків, тобто листків наступного ярусу. Прикладом першого типу листків можуть бути листкові пластинки веху: Sium latifolium, S. sisaroideum (Петрова, Барыкина, 2005) та ін.; другого типу - листки Campanula rotundifolia, Triphyophyllum peltatum, опис яких наведено нижче.
У суходільного екотипу Sium latifolium, який зростає на відстані 5-20 м від берега, форма листкових пластинок різниться. Рослини цього виду характеризувалися гетерофілією. Листки були двох типів (рис. 1.3.6, а-в, див. вставку VI): з цілісною пластинкою (рис. 1.3.6, а, б) і розсіченою (рис. 1.3.6, а, в). Перші три листки з цілісною листковою пластинкою мали округлояйцеподібну й загостренояйцеподібну форму з серцеподібною основою, городчатим типом краю листка; середній розмір довгої осі пластинки становив 2,5 ± 0,3 см, короткої - 2,0 ± 0,2 см. Кожна особина S. latifolium у травні мала три цілісні листки й два - пірчасторозсічені, останні складалися з цілісних видовженоовальних листочків. Пірчасторозсічені листки мали 4-5 пар та одну непарну частку з городчатими краями, середній розмір довгої осі часточки становив 10 ± 1,2, короткої - 2,9 ± 0,5 см. Парні листкові часточки розміщені супротивно.
Типовим прикладом ускладнення форми листкової пластинки є листки трави Senecio lautus (Asteraceae), яка зростає у тріщинах скель Нової Зеландії (Burns, 2005). Перші листки цього виду мали цільну видовжену литкову пластинку з коротким черешком (рис. 1.3.7; 8-ий і 9-ий листки зліва), при дозріванні трави форма наступних, заново сформованих листків сильно змінювалася в порівнянні з ювенільними листками: у них поступово утворювалося парне число виступів - зубців (від 4 до 10 мм завдовжки), розмір пластинки збільшується майже удвічі (рис. 1.3.7).

Рис. 1.3.7. Листки різної форми, зібрані з однієї рослини Senecio lautus на південному побережжі Велінгтона, Нова Зеландія. Зліва направо: поступовий ряд дорослих розсічених листків до перших загостренояйцевидних ювенільних (справа) (Burns, 2005)
У рослин Pastinaca sativa, які розвиваються в умовах відкритого грунту, закладка та розвиток прикореневої розетки відбувається, як і в Apium graveolens (Наумов, 2009). Нижні пари листочків прикореневої розетки зазнають змін, пов'язаних з тим, що ці пари стають розсіченими (рис. 1.3.8; див. вставку УІІ). Особливо чітко цей процес виявляється на другій- четвертій парі бічних листочків при основі листка.
У Campanula rotundifolia прикореневі листки загостренояйцевидні зі зубчатим краєм (рис. 1.3.9, а; див. вставку УІІ), черешки довгі. Розеточні листки утворюються рано весною, і зростають у затінку оточуючих рослин, тоді як в червні на стеблі формуються лінійні листки (рис. 1.3.9, б, в).
Якщо рослина зростає на слабкому освітленні, то на стеблі замість видовжених листків будуть формуватися округлі або ж серцеподібної форми листки (Барабанов, 2006).
Для рослини Triphyophyllum peltatum, які зростають в тропіках Африки, характерна наявність сезонної гетерофілії (Green et al., 1979), що виявляється в трьох типах листків. Перший тип - це списоподібні видовжені листки прикореневої розетки, їхній розмір 35,5 х 5 см, такі листки характерні для ювенільної фази рослини, у якої пагін не перевищує 50 см у висоту (рис. 1.3.10, а; див. вставку VE). Коли рослина починає квітнути, пагін витягується до 50 м, на ньому з'являються лінійні комахоїдні листки із залозками (другий тип листків) (рис. 1.3.10, б); такі листки маленькі, вусикоподібні (діаметром до 5-8 мм), часто без листкової пластинки, за допомогою залоз листки перетравлюють комах і абсорбують поживні речовини після розчинення комахи-жертви. Другий тип листків інколи може мати базальну листкову пластинку. Після цвітіння на пагоні утворюється й третій тип листків лопатчатої форми - стеблові листки з двома гачкоподібними виростами на кінчику пластинки (рис. 1.3.10, в), за допомогою яких вони чіпляються за стовбури дерев, середній розмір третього типу листків становить 17,8 х 3,7 см (Green et al.,1979; Albert et al., 1992; Bringmann et al., 2002).
Для ендемічних рослин з роду Cynea (Campanulaceae: Lobelioidae), що зростають на океанічних островах в сильно затінених місцях на Гавайському архіпелазі, також характерне явище гетерофілії: нижні листки розсічені, мають форму S-літери, колючкоподібні, верхні листки мають цільну пластинку (Givnish, 1987; Givnish et al., 1994; Lee etal., 1990). Ці дослідники вважають, що нижні листки завдяки своїй формі можуть сприймати слабкі сонячні промені в сильно затінених місцях їхнього зростання. Автори, вивчаючи наявність гетерофілії у 55 видів роду Cynea, встановили, що це явище характерне для 18 видів, які проходять стадію формування молодих гачкоподібних листків лише на деяких островах цього архіпелагу. Згідно думки цих авторів, гетерофілія ендемічних рослин на океанічних островах виникла в процесі еволюції як один із засобів захисту рослин (утворення листків «колючок») від механічних пошкоджень травоїдними (Givnish, 1987; Givnish et al., 1994).