БОТАНІКА 3 ОСНОВАМИ ГІДРОБОТАНІКИ - 2010
11. ОСНОВИ ГІДРОЕКОЛОГІЇ РОСЛИН І ФІТОЦЕНОЛОГІЇ
Вивчення рослин і тварин свідчить, що в однотипних умовах існування в них виникають схожі пристосування незалежно від їх систематичної спорідненості. Саме екологічні фактори визначають характер та особливості тих чи інших адаптацій організмів.
Різноманітність екологічних умов і відповідно різноманітність адаптацій організмів стало об'єктивною передумовою створення значної кількості різних екологічних класифікацій і виділення численних екологічних груп організмів. Серед усієї різноманітності екологічних факторів досить важко виділити найважливіші для класифікації. Крім того, використовуючи який-небудь один фактор, неможливо відобразити всі сторони пристосованості організмів до середовища.
Екологічна група - це сукупність організмів різних видів незалежно від їх систематичної належності, що характеризуються подібними адаптаційними особливостями до конкретного екологічного фактора. Екологічна група виділяється, як правило, на основі відношення різних організмів до якогось одного фактора; адаптації організмів до дії комплексу різних екологічних факторів знаходять своє вираження в життєвій формі або екоморфі.
Екологічні групи рослин можна виділяти за відношенням до різних екологічних факторів.
11.1. Класифікація екологічних факторів
У широкому розумінні оточуюче середовище (довкілля) - це сукупність матеріальних тіл, явищ і енергії, які впливають на живий організм. Впливають ці елементи по-різному. Деякі з них практично байдужі для організму рослин (наприклад інертні гази у повітрі). Інші чинять на рослини істотний вплив - це екологічні фактори.
За традиційною (класичною) екологічною класифікацією фактори, які впливають на розвиток усіх живих організмів і рослин у тому числі, розділяють на абіотичні, не пов’язані з життєдіяльністю організмів, біотичні, обумовлені такою діяльністю, і антропогенні, обумовлені діяльністю людини. Одні види легко переносять значні коливання тих або інших факторів середовища, інші, навпаки, можуть існувати в межах невеликих їх змін. Багато чинників, особливо абіотичних, є лімітуючими, тобто тими, які обмежують існування гідробіонтів.
Абіотичні фактори розділяються на декілька груп:
Кліматичні: світло, температура, повітря (його склад і рухи), вологість грунту і повітря, а також опади.
Едафічні (ґрунтові) - механічний і хімічний склад ґрунтів, їх фізичні властивості і ін.
Топографічні (або орографічні) - вплив особливостей рельєфу на рослинність.
Хімічні фактори (мінералізація води і т.д.)
Всі ці чинники неживої природи безпосередньо або опосередковано впливають на наземні рослини. Водні рослини зазнають впливу іншого комплексу факторів водного середовища.
Біотичні фактори - це форми впливу живих організмів один на одного. Кожен організм постійно відчуває на собі прямий чи опосередкований вплив інших істот, вступає у зв’язки з представниками свого виду і інших видів - рослинами, тваринами, мікроорганізмами, залежить від них і сам здійснює на них вплив.
Антропогенні (антропічні) фактори - це форми діяльності людського суспільства, які призводять до зміни природних екосистем як середовища існування інших видів або безпосередньо відбиваються на їх житті.
У ході розвитку людського суспільства спочатку мисливство, а потім сільське господарство, промисловість і транспорт дуже змінили природу планети. Антропогенний вплив на біоту Землі продовжує стрімко зростати.
Реакція рослин на вплив середовища. На несприятливий вплив зовнішнього середовища живий організм реагує або певною дією - униканням несприятливих впливів (наприклад, зміною поведінки), або зміною стану і фізіологічних функцій - набуття витривалості (толерантності) до негативних чинників. Ці реакції забезпечують виживання виду у несприятливих умовах.
Уникання несприятливих умов більш властиве тваринам, які переміщуються з несприятливого середовища у сприятливе (таксиси, міграції тварин тощо). Вищі рослини можуть переміщувати частини тіла у сприятливіші умови завдяки ростовим реакціям і повільним змінам положення певних частин тіла (наприклад повертанням листків перпендикулярно до напрямку падаючого проміння). Нижчі рослини (водорості) у несприятливих умовах можуть утворювати
спори, які мають міцну оболонку і дуже легкі, можуть переноситися вітром і течіями.
Отже, основна «стратегія життя» рослин - це пластичність структур і функцій, пристосування будови і процесів життєдіяльності до навколишнього середовища.
Ці зміни можуть бути не спадковими - модифікаційна мінливість, або спадковими - змінами генотипу.
Морфологічні адаптаційні зміни можна побачити на різних рівнях організації - від клітинного до рівня цілого організму (зміна розміру, форми, співвідношення різних органів і ін.)
Фізіологічна і біохімічна пластичність проявляється у зміні хімічного складу, інтенсивності і стійкості фізіологічних процесів.
Адаптація проявляється у динамічній відповідності морфофізі-ологічної організації і пристосувальних реакцій тварини або рослини до типових і провідних умов середовища, в яких організм формувався. Адаптивна організація забезпечує не лише виживання індивідууму, але й успішне існування виду.
Вплив рослини на середовище. Усі рослини певним чином впливають на навколишнє середовище. Маленькі накипні лишайники на скелях виділяють органічні кислоти у процесі життєдіяльності і таким чином сприяють біологічному вивітрюванню коєної неорганічної речовини. Під кроною дерева формується особливий мікроклімат - тінь, нижча температура, інший вміст газів і летких речовин. Ще більший вплив на довкілля мають великі рослинні угруповання. Вони не лише впливають на середовище, але і на інші живі організми і самі зазнають впливу.
Отже, розрізняють два поняття місцезростання рослини і комплексу діючих на неї факторів: екотоп - первинний комплекс факторів фізико-географічного середовища; біотоп - умови середовища, видозмінені життєдіяльністю живих організмів.
Наприклад, зарості вищих водних рослин негативно впливають на проникнення сонячної радіації в товщу води. Так, у порівнянні з відкритими акваторіями, на поверхню води з рогозом широколистим (50-60 % проективного вкриття) потрапляє 40 %, а з очеретом звичайним (90-95 % проективного вкриття) - тільки 25 % сонячної радіації. Ще менше сонячної енергії проникає в товщу води. Наприклад, на глибину 0,3 м в заростях лепешняка плаваючого із покриттям 90 % поступає 15 %, а при вкритті 60 % - 80 % сонячній енергії.
Поглинання у верхніх шарах води значної частини сонячної радіації різко обмежує розповсюдження в товщі води фотосинтезуючих рослин. Вони можуть розвиватися на відносно невеликій глибині в континентальних водоймах, морях і океанах.
11.2. Екологічні фактори і екологічні групи рослин
Вода як екологічний фактор. Живі організми не можуть існувати без води, вона є їх невід'ємною частиною, займаючи від 40 до 90% об'єму рослин. Вода необхідна для фотосинтезу, транспірації, ферментативної активності, поглинання і переміщення ґрунтового розчину, пластичних речовин, дихання, запліднення. Велику частину рослинної клітини складає вода. Цитоплазма в середньому містить 85—90 % води, і навіть такі багаті ліпідами клітинні органоїди, як хлоропласти і мітохондрії, містять не менше 50 % води. Вода в рослинній клітині існує в двох формах: конституційна вода, зв'язана водневими зв'язками із структурами макромолекул, і резервна вода, не зв'язана, яка міститься у вакуолях. У резервній воді зазвичай розчинені вуглеводи, різні органічні кислоти і т. п., унаслідок чого вона може брати участь в стабілізації внутріклітинного осмотичного тиску.
У різних точках земної кулі співвідношення між надходженням вологи і її витратами нерівномірне. В полярних областях, пустелях, тропіках, поблизу океанів випаровування води досягає великих розмірів. Області, де випаровування перевищує річну суму опадів, а рослини відчувають нестачу вологи, називають аридними, а області, де рослини забезпечені вологою, — гумідними.
За відношенням до води виділяють такі екологічні групи рослин (гідроморфи): ксерофіти, мезофіти, гігрофіти, гідрофіти, гідатофіти. Між ними можуть бути перехідні форми.
Ксерофіти (гр. xeгos — сухий) — рослини, що зростають в місцях з недостатнім зволоженням і мають пристосування, які дозволяють їм добувати воду при її нестачі, обмежувати випаровування води або запасати її на період посухи. Ксерофіти краще, ніж інші групи рослин, здатні регулювати водний обмін і тому під час тривалої посухи залишаються в активному стані. Вони поділяються на два основні типи — сукуленти та склерофіти. Сукуленти — це м'ясисті рослини з сильно розвинутою водозапасаючою паренхімою в різних органах. їх коренева система неглибока, але сильно розпрос-
терта. Прикладами сукулентів є: кактуси, кактусовидні молочаї, алое, агави, молодило, очиток, аспарагус. Склерофіти — це сухі на вид рослини, часто з вузькими і дрібними листками, які покриті волосками або восковим нальотом. Вони мають добре розвинуту склеренхіму, тому без значних наслідків можуть втрачати до 25% води не в'янучи. Прикладами склерофітів є ковила, тонконіг вузьколистий, костриця овеча, полини та деякі інші рослини).
Мезофіти (гр. mesos — середній) — рослини, що потребують для свого розвитку помірного грунтово-повітряного зволоження. Це найчисленніша екологічна група, що об'єднує рослини, які можуть переносити нетривалу і не дуже сильну засуху. Це рослини, що зростають при середньому зволоженні, помірному тепловому режимі і досить хорошому забезпечені мінеральним живленням. Вони мають добре розвинену кореневу систему, швидко ростуть за умов достатньої родючості грунту. Тканини листка мезофітів мають стовпчасту і губчату паренхіму, розвинену сітку жилок, середнього розміру міжклітинники.
За здатністю регулювати свій водний обмін одні з цих рослин подібні до гігрофітів, інші — до посухостійких форм. До цієї групи можна віднести вічнозелені дерева верхніх ярусів тропічних лісів, листопадні дерева саван, літньозелені листяні породи лісів помірних широт, кущі підліску, трав'янисті рослини широкотрав'я, рослини заплавних і не дуже сухих суходільних лук, пустельні ефемери та ефемероїди, багато бур'янових і більшість культурних рослин.
Гігрофіти (гр. hydros - вологий) — рослини вологих місцезростань: боліт, берегів рік та озер, вологих луків і лісів. Вони не переносять дефіціту вологи і не пристосовані до її обмеження. Для них характерним є розвиток гідатод, або водяних продихів, що виділяють краплиннорідку воду. їх листки часто тонкі, слабо розвинута кутикула, обводненість тканин — до 80%. Найтиповішими гігрофітами є трав'янисті рослини і епіфіти вологих тропічних лісів: епі-фітні види папоротей, болотні пальми, папірус та інші рослини, а в наших широтах калюжниця, вовче тіло болотне, болотні осоки, очерет, рогіз та багато інших рослин. Із культурних рослин до гігрофітів належить рис. Характерними особливостями гігрофітів є великі листки з добре розвиненими міжклітинниками, розміщення коренів у поверхневих горизонтах грунту, розвиток так званих дихальних коренів. Структура листків і стебел, характерна для гігрофітів, на-
зиваєгься гігроморфною. Ця їх структура разом із слабкою продиховою регуляцією транспірації спричинює швидке в'янення гігрофітів при зменшенні вологості повітря і грунту.
Між типовими гігро- і мезофітами є перехідні групи. Це переважно болотні і лучні злаки і осоки, їх відносять до гігромезофітів. Болотяні травянисті рослини називають також гелофітами (рис. 11.1).
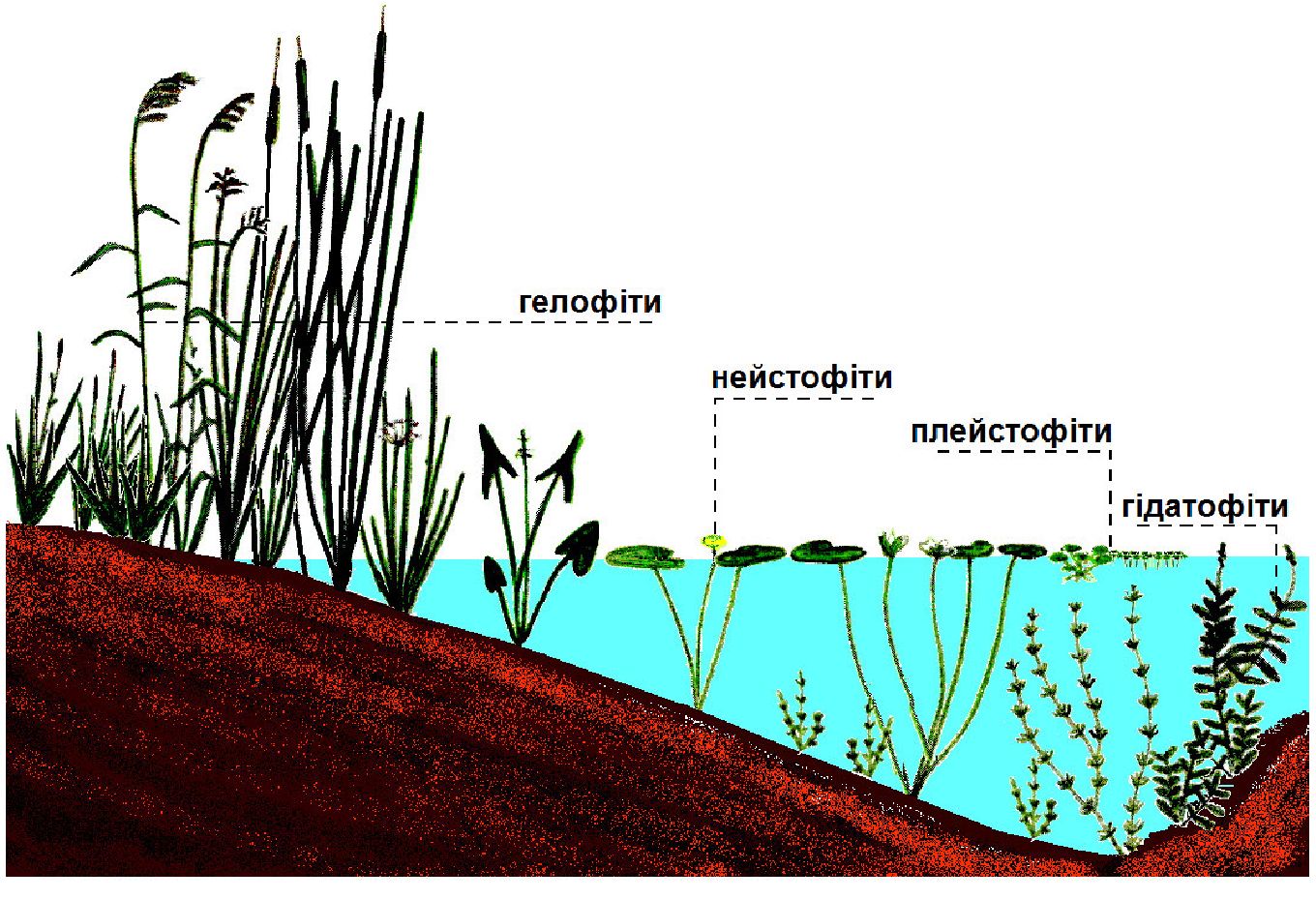
Рис. 11.1. Екологічні групи водних рослин
Гідрофіти (гр. hydro — вода) — це наземно-водні рослини, що частково занурені у воду і зростають вздовж берегів, на мілководді, на болотах. У них краще, ніж в гідатофітів, розвинуті провідні та механічні тканини, добре виражена аеренхіма, досить висока інтенсивність транспірації. До цієї групи відносяться такі рослини, як очерет звичайний, калюжниця болотна, частуха подорожникова, бобівник трилистий та інші види.
Гідатофіти (гр. hidatos — вода, волога + гр. phyton — рослина) — види рослин, що повністю або значною мірою занурені у воду. До групи гідатофітів відносяться такі рослини, як елодея канадська, рдесники, кушир, водяний жовтець, жабурник. Рослини, які повністю занурені у воду, а над поверхнею підносяться лише генеративні органи, називають еугідатофітами (рдесники, водопериця, елодея).
Серед гідатофітів є рослини листя яких плаває на поверхні води. Частина з них вільно плаває на поверхні води - плейстофіти (ряска мала, спіродела багатокоренева), а інші укорінені - нейстофіти (латаття біле, глечики жовті).
Водне середовище має ряд особливостей, до яких у рослин виробились різноманітні пристосування (редукція продихів, добре розвинута аеренхіма, відсутність кутикули тощо). Вищі водні рослини є вторинноводними організмами - наземними рослинами, що пристосувалися до життя у воді. їх види належать до дуже різних і віддалених одна від одної родин.
До основних пристосувань вищих рослин до водного середовища існування відносяться такі:
1) переважання вегетативного розмноження;
2) посилений ріст, порівняно з наземними рослинами;
3) недорозвинутість або відсутність деревини в стеблах;
4) редукція кореневої системи або зміна її функцій;
5) порівняно велика площа поверхні тіла для кращого газообміну, відсутність диференціації паренхіми на палісадну і губчасту;
6) гетерофілія;
7) добре розвинута аеренхіма
8) виділення слизу спеціальними залозками, що запобігає вимиванню поживних речовин з рослини;
9) переважна більшість ВВР є багаторічниками.
У гідатофітів дуже поширена гетерофілія (різнолистість). Листки, що плавають на поверхні води, цілісні й більші за розміром, а занурені у воду — розсічені на дрібні частки, їх епідерміс дуже тонкий, позбавлений кутикули, пластинка тонка, без продихів (рис. 11.2). Плаваючі листки, навпаки, мають складнішу будову, епідерміс верхнього боку листка має кутикулу, велику кількість продихів, а мезофіл диференційований на стовпчасту і губчату паренхіми.
Коренева система у плаваючих рослин розвинена слабко, листки мають гідатоди.
Водні рослини відрізняються за розміром, і за цією ознакою виділяють дві групи - мікрофіти і макрофіти. Мікрофіти - це мікроскопічні водні рослини, водорості. Відповідно, макрофіти - це видимі неозброєним оком водні рослини. Макрофітами називають вищі водні рослини і нижчі рослини - макроскопічні водорості (харові, зелені тощо).
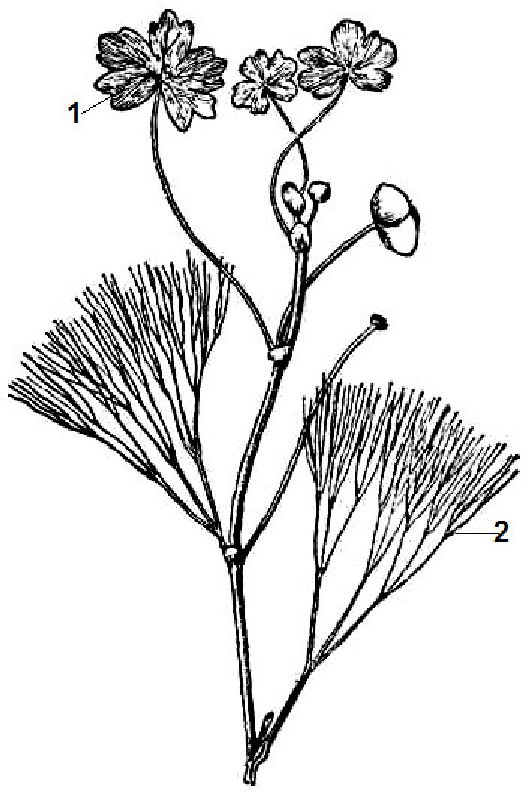
Рис. 11. 2. Гетерофілія у водяного жовтеця: 1 — підводне листя; 2 — надводне листя
У нижчих рослин (водоростей) тіло не розчленоване на органи і тканини, на відміну від вищих, у яких є складно побудовані вегетативні і генеративні органи: стебло, листок, корінь, квітка.
Для більшості водоростей вода — постійне місце існування, проте багато водоростей можуть жити і поза водою. За стійкістю до висихання, серед рослин, що мешкають на суші, виділяють пойкілогідричні — не здатні підтримувати постійний вміст води в тканинах, і гомойогідричні — здатні підтримувати постійну гідратацію тканин. У пойкіло-гідричних рослин (синьозелені і деякі зелені водорості) клітини при висиханні стискаються без необоротної зміни ультраструктури і, отже, не втрачають життєздатності. При зволоженні вони відновлюють нормальний метаболізм. Мінімальна вологість, при якій можлива нормальна життєдіяльність таких рослин, різна. її значення зумовлює, зокрема, розповсюдження аерофітів (повітряних водоростей).
Для гомойогідричних рослин обов'язкова наявність крупної центральної вакуолі, за допомогою якої стабілізується водний запас клітини. Проте клітини з крупними вакуолями в значній мірі втрачають здатність до висихання. До гомойогідричних водоростей відносяться, наприклад, деякі повітряні зелені і жовтозелені водорості (иіойтсйа^, Уаисйегіа^), що зазвичай поселяються в умовах постійної надмірної зволоженості.
Рух води. Величезну роль в житті водних рослин відіграє рух води. Абсолютно стоячої, нерухомої води не існує, отже, практично всі водні рослини — мешканці текучих вод. У будь-яких континентальних і морських водоймах спостерігається відносний рух водних мас, що забезпечує притік поживних речовин і видалення продуктів життєдіяльності рослин.
Швидкість течії - фактор, який за своєю значимістю прирівнюється до впливу антропогенного забруднення водойм. Thienemann A. (1912) році запропонував поділити водні організми за відношенням до швидкості течії на три групи: реофіли, реобіонти та реоксе-ни. До групи реофілів, здатних рости при швидкості течії від 0,7 до 1,2 м/ с, відносяться лепешняк плаваючий (Glyceria fluitans), очерет звичайний (Phragmites australis), сусак зонтичний (Butomus umbellatus), рдесник злаколистий (Potamogeton gramineus), рдесник пронизанолистий (Potamogeton perfoliatus), плавун щитолистий (Nymphoides peltata). Більшість занурених водних рослин є реобіо-нтами, ростуть в проточних водах (від 0,2 до 0,7 м/ с). У стоячих та малопроточних водах переважно зустрічаються реоксени - лепеха звичайна (Acorus calamus), калюжниця болотна (Caltha palustris), плавушник болотний (Hottonia palustris).
На ділянках із швидкою течією знижується видова різноманітність до одного-двох пристосованих до таких умов рослин та проективне вкриття до 10-15%. На ділянках з повільною течією зростає гострота конкуренції між водними рослинами, адже тут значно ширший видовий спектр та високе проективне вкриття (до 100%).
Донні водорості, що ростуть в умовах проточності, мають переваги в порівнянні з водоростями, що ростуть в малорухливих водах. Один і той же рівень фотосинтезу може бути досягнутий рослинами в умовах течії при меншій освітленості, що сприяє росту більших за розмірами водоростей з великим вмістом поживних речовин. Рух води запобігає осіданню на скелях і каменях мулистих частинок, які заважають закріпленню зачатків водоростей, змиває з поверхні грунту тварин, що харчуються водоростями. Хоча при сильній течії або сильному прибої відбувається пошкодження водоростей або відривання їх від грунту, рух води перешкоджає поселенню мікроводо-ростей і мікроскопічних стадій макроводоростей. Як правило, місця з інтенсивним рухом води відрізняються значним розвитком донних водоростей.
В Арктиці багаторічні бурі водорості (Fucus, Laminaria) найлегше знайти біля берега серед валунів і виступів скель, що перешкоджають руху льоду, який може знищити зарості рослин.
Світло. Основне джерело енергії у біосфері - сонячне випромінювання. Енергію світла засвоюють фототрофні організми (вищі і нижчі рослини), перетворюючи її на доступну для консументів форму - енергію хімічних зв’язків органічних речовин.
Світло необхідне для пересування поживних речовин у рослині, здійснення транспірації та якісних біохімічних перетворень. Енергія сонячного променя витрачається на здійснення морфогенетичних процесів: проростання насіння, закладення і розвиток вузла кущіння у злаків, росту проростків, формування стебел, розвиток квітки, визрівання плодів тощо. Всі ці процеси потребують певної кількості й тривалості освітлення.
Сонячна радіація відіграє виключно важливу роль у функціонуванні водних екосистем. З нею пов'язана поведінка і розселення гідробіонтів у біотопі. Серед гідробіонтів є організми, що інтенсивно розвиваються у верхніх шарах води, куди поступає найбільша кількість сонячної енергії. Це переважно автотрофні організми: водорості, фотосинтетичні бактерії, вищі водні рослини. В процесі фотосинтезу вони запасають велику кількість енергії у вигляді органічної речовини (первинній продукції), яка потім використовується організмами інших трофічних рівнів.
Автотрофні водні організми (водорості, вищі водні рослини) використовують спектр сонячної радіації в діапазоні 380-710 нм. Саме така радіація найадекватніше впливає на фізіологічні процеси, пов'язані з фотосинтезом водних рослин. Ця область спектру називається фотосинтетично активною радіацією (ФАР). Пряма сонячна радіація містить 28-43 % ФАР, а розсіяна - 50-60 %.
Світловий промінь, що падає на водну поверхню, крім віддзеркалення і заломлення зазнає дифракції, поляризації і спектрального розщеплення. Крім того, він поглинається під час проходження крізь товщу води і відбивається від завислих у воді частинок, унаслідок чого на різні горизонти доводиться різна кількість сонячної енергії, а це обумовлює зниження освітленості з глибиною.
Важливою з екологічного погляду властивістю води є здатність пропускати сонячне світло. Вона залежить від кольору води і її прозорості. Остання визначається молекулярною структурою і концентрацією розчинених органічних, переважно забарвлених (гумінові кислоти, фульвокислоти і т. п.), речовин, завислих частинок і планктонних організмів.
Верхній шар води, в який проникає достатня кількість світла для синтезу рослинами органічної речовини з використанням сонячної енергії, називається фотичним, нижній шар, куди не поступає соня
чна енергія, - афотичним. Зона проникнення світла, в якій інтенсивність фотосинтезу перевищує інтенсивність дихання рослин, має назву евфотичної зони. її нижня межа, де фотосинтез врівноважує інтенсивність дихання, називається компенсаційним горизонтом.
У воді найінтенсивніше поглинаються довгі хвилі сонячної радіації, яка найбільш важлива для фотобіологічних процесів. Крізь товщу прісних і морських вод проникає переважно випромінювання блакитної частини спектру з довжиною хвилі 475-480 нм. У процесі фотобіологічних реакцій енергія сонячної радіації поглинається дискретними частинками, що називаються фотонами, або квантами. Фотосинтез у бактерій протікає в спектральному діапазоні 400-900 нм, вищих зелених рослин - 400-700 нм, водоростей - 400660 нм. Якщо хвилі коротше 300 нм, порушується молекулярна структура білків і нуклеїнових кислот і відповідно — нормальне функціонування живих систем. От чому представляють загрозу для біосфери зменшення і розриви озонового шару, який затримує проникнення на Землю саме таких квантів сонячного випромінювання.
За відношенням до світла виділяють декілька екологічних груп рослин, або геліоморф. геліофіти (гр. helios — сонце + гр. phyton — рослина) — світлолюбні рослини, що віддають перевагу яскраво освітленим місцям зростання; сціофіти (гр. scia — тінь) — тіневитривалі рослини, що здатні переносити значне затінення; геліосціо-фіти — рослини, які добре ростуть на освітлених місцях, але можуть переносити більше чи менше затінення.
Світлолюбні рослини, або геліофіти, з оптимальним розвитком при повному освітленні. Ці види погано переносять навіть незначне затінення - лепеха звичайна (Acorns calamus), частуха подорожни-кова (Alisma plantago-aquatica), сусак зонтичний (Butomus umbellatus), півники водяні (Iris pseudacorus), лотос горіхоносний (Nelumbo nucifera), глечики жовті (Nuphar lutea), латаття біле (Nymphaea alba), рдесники блискучий (Potamogeton lucens) і прони-занолистий (P. perfoliatus), спіродела багатокоренева (Spirodela polyrrhiza).
Тіньові рослини, або сціофіти, з оптимальним розвитком в межах від 1/10 до 1/3 повного освітлення. З водних рослин сіофітами є образки болотні (Calla palustris), кушир занурений (Ceratophyllum demersum), хвощ річковий (Equisetum fluviatile), молодильник озерний (Isoëtes lacustris), м’ята водяна (Mentha aquatica), пухирник звичайний (Utricularia vulgaris).
Тіньовитривалі рослини, або геліосціофіти, різного ступеня. До цього типу належать види, що краще ростуть при освітленні, але здатні переносити затінення без особливого пригнічення - альдро-ванда пухирчаста (Aldrovanda vesiculosa), меч-трава болотна (Cladium mariscus), вовче тіло болотне (Comarum palustre), елодея канадська (Elodea canadensis), лепешняк великий (Glyceria maxima), ряска триборозенчаста (Lemna trisulca), рдесник плаваючий (Potamogeton natans), сальвінія плаваюча (Salvinia natans).
Водорості, особливо мікроскопічні, характеризуються найвищим ККД засвоєння світлової енергії у порівнянні з іншими фотосинте-зуючими організмами. Багато видів здатні до міксотрофізму і ефективної утилізації світла низької інтенсивності.
Водорості по відношенню до світла поділяють на дві групи: геліофільні і геліофобні.
Геліофільні (світлолюбні) водорості потребують значної кількості світла для нормальної життєдіяльності і фотосинтезу. До них відносяться більшість синьозелених і значна кількість зелених водоростей, що масово розвиваються в літній час у поверхневих шарах води.
Геліофобні водорості пристосовані до умов низької освітленості. Наприклад, більшість діатомових водоростей уникають яскраво освітленого поверхневого шару води і в малопрозорих водах озер інтенсивно розвивається на глибині 2-3 м, а в прозорих водах морів — на глибині 10-15 м.
У водоростей різних відділів залежно від складу пігментів — фоторецепторів, максимальна інтенсивність фотосинтезу спостерігається при різній довжині світлових хвиль. У наземних умовах якісні характеристики світла досить постійні, так само як інтенсивність фотосинтезу. При проходженні крізь воду світло червоної і синьої області спектру поглинається і на глибину проникає зеленувате світло, що слабо сприймається хлорофілом. Тому там виживають в основному червоні і бурі водорості, що мають додаткові фо-тосинтезуючі пігменти (фікоціани, фікоеритрини тощо), здатні використовувати енергію зеленого світла. Це зумовлює вертикальний розподіл водоростей в морях і океанах: у приповерхневих шарах, як правило, переважають зелені водорості, глибше — бурі, на найбільш глибоководних ділянках — червоні. Проте подібна закономі
рність не є абсолютною. Багато водоростей здатні існувати в умовах крайньої низької, не властивої їм, освітленості, а іноді і в повній темряві. При цьому у них можуть відбуватися певні зміни в пігментному складі або в способі живлення. Так, у синьозелених водоростей в умовах низької освітленості пігментний склад може змінюватися у бік переважання фікобілінів, колір трихомів при цьому змінюється від синьозеленого до пурпурного. Представники багатьох відділів водоростей (наприклад, евгленові) здатні за відсутності світла і надлишку органічних речовин переходити до сапротрофного способу живлення.
Лише 10% морської води має достатні світлові умови для фотосинтезу. 90% води знаходиться в абсолютній темряві й отримує поживні речовини передусім завдяки опусканню їх із евфотичної зони. Ультрафіолетове та довгохвильове світло проникає на меншу глибину, ніж синьо-зелене. Рослини, що живуть близько від поверхні, наприклад Шяа sp., абсорбують частину світла подібно до наземних рослин. Червоні водорості використовують жовто-зелене світло, яке глибоко проникає в воду, за допомогою спеціальних пігментів, які переносять енергію на хлорофіл а.
Абсорбція бактеріальних пігментів проникає далеко в невидимий діапазон світлового випромінювання (рис. 11.3). Продуценти можуть існувати в морських екосистемах завдяки пристосуванню абсорбційної системи до різних світлових зон на глибині до 200 м.
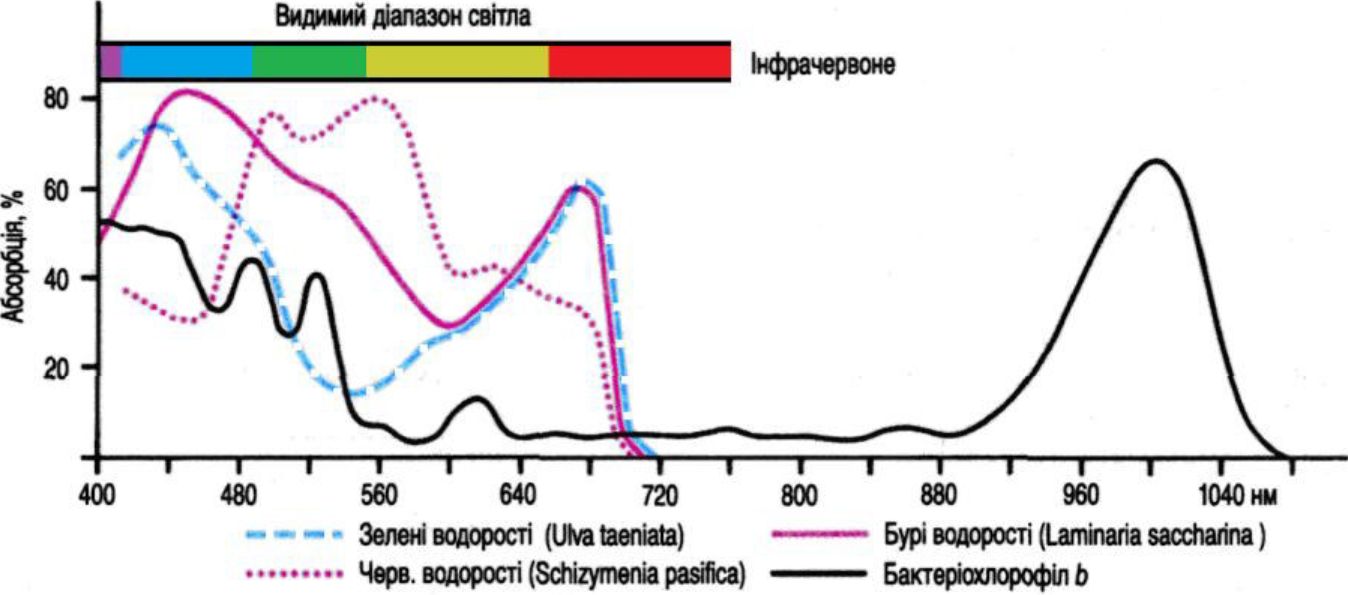
Рис. 11. 3. Абсорбція світла водоростями і бактеріальними пігментами
Інші процеси, що протікають за участю сонячної енергії, не по-
в'язані безпосередньо з перетворенням сонячного світла в енергію хімічних сполук. Світло може виступати як інформативний чинник, що визначає фотоперіодичні реакції рослин, що синхронізують етапи їх репродуктивного циклу, здійснюються за допомогою пігменту фітохрому.
Фототаксис - це рух організмів під впливом одностороннього освітлення. У водоростей, найпростіших, деяких багатоклітинних організмів він виражається у переміщенні в більш освітлені ділянки водойм. Найбільш активний фототаксис викликають ультрафіолетові, фіолетові і сині промені сонячного спектру.
Фототропізм виявляється в зміні напряму росту гідробіонтів у відповідь на односторонню дію світла.
Температура. Водні екосистеми характеризуються повільним охолоджуванням і нагріванням води внаслідок надзвичайно високої її теплоємності. Це обумовлено витрачанням певної частини теплової енергії на розрив водневих зв'язків в асоційованих молекулах. При значному підвищенні температури повітря вода стає теплішою, але її температура ніколи не досягає атмосферної через високу теплоту пароутворення. У спекотні дні зростає інтенсивність випаровування води, а відповідно і віддача тепла, що запобігає її перегріву. І навпаки, при зниженні температури нижче 0°С і утворенні льоду вивільняється значна кількість тепла, і тому вода охолоджується поволі. Завдяки високій теплоємності води діапазон коливань температури водного середовища, в якому живуть гідробіонти, рідко перевищує 1-35 °С. У порівнянні з гідробіонтами наземні організми вимушені пристосовуватися до значно більшого діапазону коливань температури навколишнього середовища.
Екологічне значення температури у першу чергу проявляється через вплив на розподіл гідробіонтів у водоймах і на швидкість протікання різних життєвих процесів. Види, які живуть в широкому температурному діапазоні, називаються евритермними (наприклад, зелені водорості з порядку едогонієвих, стерильні нитки яких можна знайти в дрібних водоймах з ранньої весни до пізньої осені), а у вузькому - стенотермними. До стенотермних відносяться, наприклад, кріофільні (холодолюбні) водорості, що ростуть тільки при температурах, близьких до точки замерзання води. Серед кріофіль-них водоростей переважають зелені, синьозелені і діатомові. Розвиваючись у масовій кількості, вони можуть викликати зелене, жовте,
блакитне, червоне, коричневе, буре або чорне «цвітіння» снігу або льоду. Зелене забарвлення снігу викликає Raphidonema nivale, червоне — Chlamydomonas nivalis, коричневе — Ancylonema nordenskioeldii. Ці водорості знаходяться в поверхневих шарах снігу або льоду та інтенсивно розмножуються при температурі близько 0°С. Лише небагато з них мають стадії спокою, більшість позбавлені будь-яких спеціальних морфологічних пристосувань для переживання низьких температур.
Ці види об'єднує здатність витримувати замерзання без порушення клітинних структур і потім, при відтаненні, швидко відновлювати вегетацію, використовуючи мінімальну кількість тепла.
Загалом, водоростям властивий, напевно, найширший діапазон температурної стійкості. Вони здатні існувати в крайніх температурних умовах — як на поверхні льоду і снігу, так і в гарячих джерелах, температура яких близька до точки кипіння води.
Термофільні водорості ростуть при температурі води 35—52 °С, а в окремих випадках до 84 °С і вище, нерідко при підвищеному вмісті мінеральних або органічних речовин (забруднені гарячі стічні води заводів, фабрик, електростанцій або атомних станцій). Типовими мешканцями гарячих вод є ціанеї, у меншій мірі — діатомові і деякі зелені водорості. Специфічно термофільних видів небагато. Більшість водоростей, виявлених в гарячих джерелах, здатні витримувати високі температури, але краще розвиваються в умовах звичайних температур, тобто по суті є мезотермними видами.
Відношення водоростей до температурного чинника впливає на їх вертикальний розподіл у водоймах. У різних водоймах і водотоках унаслідок поглинання сонячного випромінювання верхніми шарами води прогріваються тільки ці шари. Тепла вода має меншу густину, ніж холодна, а течії, що викликаються вітром, вирівнюють її густину тільки до певної глибини. З початком вегетаційного сезону, сезону інтенсивного сонячного випромінювання, в достатньо глибоких континентальних непроточних водоймах виникає дуже стійка температурна стратифікація водної товщі. У цих водоймах утворюються обмежені один від одного маси води: теплий і легкий поверхневий шар — епілімніон і під ним маса холоднішої і щільнішої води — гіполімніон. Восени вода у водоймі охолоджується і температурна стратифікація зникає. У морях і океанах також є постійний шар температурного стрибка. Водорості можуть розвиватися тільки в епілімніоні (а саме в евфотичній зоні), причому найбільш тепло-любиві і світлолюбні організми поселяються в поверхневих шарах води, що добре прогріваються.
Вплив температури на водорості, що розвиваються у водному середовищі, дуже великий. Саме температура визначає їх географічне розповсюдження. Так, види бурої водорості роду Lessonia зустрічаються тільки в межах літньої ізотерми 10 °С, види родів Laminaria, Agarum, Alaria не перетинають літньої ізотерми 20 °С, деякі види Sargassum мешкають тільки при температурі 22—23 °С (Саргасове море). В цілому, за винятком широко розповсюджених евритермних видів (наприклад, деякі Fucales), у розповсюдженні водоростей спостерігається географічна зональність: конкретні таксони морських водоростей приурочені до певних географічних поясів. Так, крупні бурі водорості (Macrocystis) домінують в північних морях. У міру просування на південь помітнішу роль починають відігравати червоні водорості, а бурі відходять на другий план.
Добре виражена географічна зональність і у морських планктонних водоростей. Для морського тропічного фітопланктону характерне значне видове багатство при дуже низькій продуктивності. У планктоні тропічних вод надзвичайно багато представлені динофі-тові і золотисті водорості. Води тропіків бідні діатомовими водоростями, які панують у північних морях.
Серед вищих водних рослин України є види-термофіли: глечики жовті (Nuphar lutea), латаття біле (Nymphaea alba), водяний хрін земноводний (Rorippa amphibia), сальвінія плаваюча (Salvinia natans), вех широколистий (Sium latifolium), водяний горіх плаваючий (Trapa natans), азола каролінська (Azolla caroliniana)/
Хімічний склад води. У воді в розчиненому стані присутні гази і мінеральні солі. Велике екологічне значення має кількість розчиненого у воді кисню. Наявність у природних водах розчиненого кисню є необхідною умовою існування більшості гідробіонтів. Кисень потрапляє у воду з повітря і виділяється водною рослинністю в процесі фотосинтезу (фотосинтетична реаерація).
Концентрація кисню в воді залежить від температури та вмісту в воді відновників (серед них проміжних речовин, що утворюються і накопичуються в результаті діяльності мікроорганізмів, які розкладають органічні рештки). Розчинність газу незначна і залежить від температури - при 0° С - 69,5, 10° С - 53,7, 20° С - 43,3 (мг/л). У
солоній воді розчинність кисню на 20% нижча, ніж у прісній. Перенасиченість киснем можна спостерігати у водних об’єктах багатих на рослинність, яка збагачує їм воду у процесі фотосинтезу.
До факторів, що ведуть до зменшення концентрації розчиненого кисню, належить і дихання живих організмів. Дихання та окиснення проходять в водоймах безперервно, посилюючись з підвищенням температури. А фотосинтез можливий лише за наявності денного світла. Крім того, проникнення кисню з атмосфери відбувається лише за відсутності льодового покриву. В результаті цих процесів існують значні сезонні та добові коливання вмісту розчиненого кисню в воді; мінімуми спостерігаються вночі та взимку на замерзаючих водних об' єктах.
Забезпечення водних рослин життєво необхідним їм киснем ускладнене. Вищі водні рослини пристосувалися до нестачі кисню у водному середовищі формуванням розвинутої системи провітрювання, яка складаються тканиною, що містить дуже великі міжклітинники - аеренхімою. Часто в ній міжклітинні простори перевищують розміри клітин. Аеренхіма складається з живих тонкостінних клітин. Атмосферне повітря проникає в рослину через продихи або сочевички органів, що перебувають над водою, і по міжклітинниках досягає клітин і тканин органів, яким не вистачає кисню, де і накопичується у великих міжклітинниках (рис. 11.4).
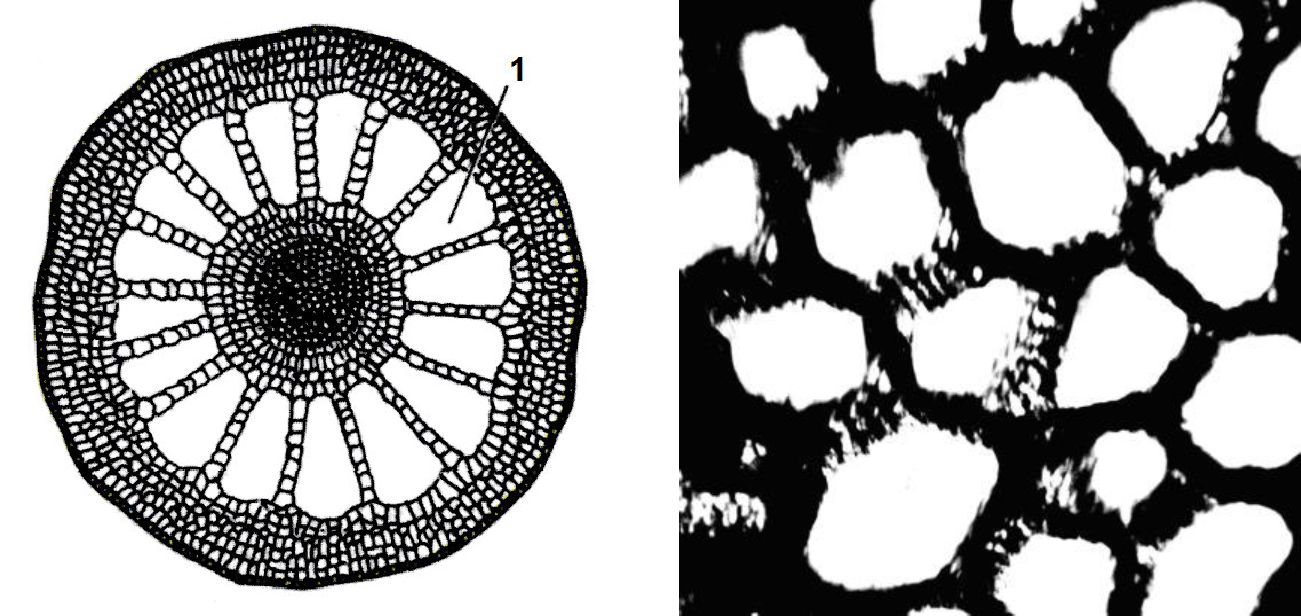
а б
Рис. 11. 4. Аеренхіма: а - зріз стебла водопериці, б - зріз листка їжачої голівки
В умовах глибоководних об’єктів, особливо Світового океану, з добре розвинутим термоклином, надлишковий кисень, який продукується фітопланктоном, виділяється в атмосферу.
Внутрішні водні об’єкти звичайно належать до числа споживачів, а не продуцентів кисню. Частково такий негативний баланс складається тому, що в умовах антропогенного тиску багато забруднюючих речовин в процесі окиснення є додатковими споживачами кисню.
Велике значення для життя рослин має вуглекислий газ. Він є умовою існування рослин і необхідний для фотосинтезу. У прісній і особливо морській воді значна кількість вуглекислого газу. Наприклад, у морській воді його міститься від 40-50 см3/л (у вільній або зв’язаній формі, що в 150 разів перевищує його концентрацію в атмосферному повітрі). Багато занурених гідрофітів здатні поглинати вуглець із води лише у формі С02, наприклад, мохоподібні, а гелофіти - з атмосфери.
Солоність і мінеральний склад води - це найважливіші чинники, що впливають на розподіл водоростей і вищих рослин.
Мінералізація морських (океанічних) вод визначається, в основному, хлоридами натрію і магнію. У одному кубічному кілометрі морської води міститься близько 35 мли т твердих речовин, зокрема 19,8 мли т хлориду натрію, 9,5 мли т магнію, 6,3 мли т сірки,
31.0 тис. т брому, 3,9 тис. т алюмінію, 79,3 т марганцю, 79,3 т міді,
11.1 т урану, 3,8 т молібдену і ін.
На відміну від океанічних і морських вод, прісні води річок, озер, водосховищ і інших водних об'єктів менш насичені розчиненими солями і містять переважно карбонати.
Залежно від солестійкості водоростей, серед них виділяють олі-гогалинні, мезогалинні, евгалинні, ультрагалинні, прісноводні і інші види. Видове багатство (чисельність видів) тісно пов'язане з солоністю води.
Практично в кожному з відділів водоростей можна знайти види, здатні мешкати в умовах крайнього засолення, і види, що живуть у водоймах з дуже низькою мінералізацією. Так, синьозелені водорості — в переважній більшості прісноводі організми, проте серед них є види, здатні розвиватися в ультрагалинних водоймах. Серед типово морських мешканців — золотистих водоростей порядку коколі-тофорид (Соссоїййорйогійа^) — зустрічаються види, поширені і в континентальних водоймах з дуже низькою мінералізацією. Діатомові водорості поширені і в морських, і в континентальних водой
мах; вони зустрічаються в умовах з різною солоністю. Проте конкретні види діатомових нерідко розвиваються тільки при певній солоності і настільки чутливі до її змін, що можуть бути використані як індикаторні організми.
Дуже чутливі до змін солоності і бурі водорості. Багато з них не може рости навіть при незначному опрісненні, тому вони бідно представлені у водах Балтійського моря з порівняно низькою солоністю. Схожу залежність від солоності водойм проявляють і червоні водорості: у Середземному морі (солоність 37—39 %о) виявлено більше 300 видів червоних водоростей, в Чорному (17—18 %о) — 129, в Каспійському (10 %о) — 22. Зелені водорості переважно прісноводні організми, лише 10 % з них зустрічаються в морях. Проте серед них є види, здатні витримувати значне засолення і навіть викликати «цвітіння» ультрагалинних водойм (наприклад, Dunaliella salina, Asteromonas gracilis).
Таким чином, водоростям в цілому властива дуже широка амплітуда солестійкості. Що стосується конкретних видів, то небагато з них здатні існувати у водоймах з різною солоністю, тобто більшість водоростей — стеногалинні види. Евригалинних видів, здатних існувати при різній солоності, порівняно небагато (наприклад, Bangia, Enteromorpha, Dunaliella).
При полімеризації високоактивних дрібних молекул в макромолекули (наприклад, при перетворенні глюкози на крохмаль) і при зворотному процесі — гідролізі високомолекулярних з'єднань, осмотичний тиск в клітині здатний швидко змінюватися. Цей механізм забезпечує стійкість окремих видів водоростей до висихання і до різких коливань солоності води.
Водорості солоних водойм — галобіонти, — ростуть при підвищеній концентрації у воді солей, що досягає 285 г/л в озерах з переважанням кухонної солі і 347 г/л в глауберових озерах. У міру збільшення солоності кількість видів водоростей зменшується; дуже високу солоність переносять лише небагато з них. У пересолених (гіпергалинних) водоймах переважають одноклітинні рухомі зелені водорості — гіпергалоби, клітини яких позбавлені оболонки і оточені лише плазмалемою (види родів Dunaliella, Asteromonas, Pedinomonas). Ці водорості відрізняються підвищеним вмістом хлориду натрію в протоплазмі, високим внутріклітинним осмотичним тиском (до 250П03 ГПа), накопиченням в клітинах каротиноїдів і
гліцерину, високою лабільністю ферментних систем і обмінних процесів. У солоних водоймах півдня України вони нерідко розвиваються в масовій кількості, викликаючи червоне або зелене «цвітіння» солоних водойм.
Більшість видів судинних макрофітів дуже чутливі до концентрації солі у воді. Але очерет звичайний, наприклад, відноситься до видів, які здатні рости в умовах значного засолення, - галофітів. Вид росте по берегах морів і заток, солених озер і інших солонува-товодних водойм, часто на околицях солончаків. Очерет звичайний росте в умовах хлоридного (до 2,5-3%) і сульфатного (до 5%) засолення. Відмирає на другий рік лише в умовах сильного хлоридного засолення (до 5%).
Екстремально галофільний вид - рдесник гребінчастий (Potamogeton pectinatus), виявлено на ділянках, де солоність складала 16-17 %.
До групи глікогалофітів відносяться руппія морська, камка морська (Zostera marina), камка мала (Zostera nana).
До галофобів відноситься плавушник болотний (Hottonia palustris).
Кислотність води. Ефект підвищеної концентрації в воді і в ґрунті іонів водню здійснює на рослини токсичний ефект, обмежуючи поглинання кореневими системами біогенних елементів. В кислому середовищі підвищується розчинність токсичних речовин. Підкислення середовища призводить до зміни гідробіоценозів, заміни видів судинних рослин на мохоподібні, що має наслідком зміну складу та властивостей грунту під рослинами, а у деяких випадках призводить до заболочування місцевості.
Кислотність води має велике значення для життєдіяльності водоростей. За відношенням до кислотності середовища виділяють види, що живуть в лужних водах — алкалофіли (базофіли) і живуть у кислих водах, при низьких значеннях pH — ацидофіли. Ацидофі-лами, наприклад, є більшість десмідієвих (Desmidiales). Найбільше видове багатство десмідієвих водоростей спостерігається в евтроф-них і мезотрофних болотах, в умовах зниженої кислотності, проте деякі десмідієві можуть зустрічатися і в лужних водах з високою мінералізацією (наприклад, Closterum acerosum, C. leibleinii). Харові, навпаки, переважно алкалофіли. їх найбільша видова різноманітність спостерігається в слаболужних водах, проте деякі з них
(Chara vulgaris) розвиваються і в кислих водах, при pH 5,0.
Більшість вищих водних рослин віддають перевагу нейтральному середовищу. До ацидофіліє належать мохи (р. Sphagnum), настурція лікарська (Nasturtium officinale), жовтець язиколистий (Ranunculus lingua), щавель прибережний (Rumex hydrolapathum), водяний різак алоевидий (Stratiotes aloides), осока пухирчаста (Carex vesicaria), бобівник трилистий (Menyanthes trifoliata), мох сфагнум, слабий ацидофіл - глечики жовті (Nuphar lutea), ситник бульбистий (Juncus bulbosus). До базифілів належать водяні жовтеці - водний (Batrachium aquatile) і закручений (B. circinatum), елодея канадська (Elodea canadensis), кушир напівзанурений (Ceratophyllum submersum), перстач гусячий (Potentilla anserina), рдесник гребінчастий (Potamogeton pectinatus) і ін. Індиферентні види можуть рости при різних значенням pH — їжача голівка мала (Sparganium minimum), рогіз вузьколистий (Typha angustifolia).
Біогенні речовини. Наявність в середовищі макро- і мікроелементів, що є необхідними компонентами тіла водних рослин, має вирішальне значення для інтенсивності їх розвитку. Концентрація мінеральних елементів у звичайній природній воді досить низька, тому коефіцієнт накопичення (відношення елементів у біоті і природній воді) складає декілька порядків.
Елементи і їх сполуки, що відносяться до макроелементів, потрібні організмам у порівняно великих кількостях. Наприклад, азот входить до складу всіх білкових молекул.
За вибагливістю до азотного живлення рослини діляться на ні-трифілів і нітрифобів.
Нітрифіли — рослини, що вимагають для свого розвитку багато легкозасвоюваних сполук азоту. Типовими нітрифілами є плавушник болотний (Hottonia palustris) і ряска триборозенчаста (Lemna trisulca), помірним нітрифілом є жабурник звичайний (Hydrocharis morsus-ranae).
Нітрифоби — рослини, що уникають багатого азотного живлення. Вони мало вибагливі до азотних сполук і розвиваються там, де цей елемент у мінімумі.
Кальцій необхідний для обміну речовин у рослині. Він нейтралізує отруйні властивості щавлевої кислоти, знижує кислотність ґрунтового розчину, чим сприяє росту рослин. Відсутність або ж нестача кальцію в ґрунті порушує фізіологічні процеси життєдіяльності
рослин, зумовлює карбонатний хлороз. Цей елемент у великих кількостях використовується морськими і прісноводими водоростями, що відкладають навколо тіла «чохли» з солей кальцію (деякі червоні і харові водорості).
За відношенням до кальцію рослини діляться на такі екологічні групи: кальцифіли, кальцифоби та індиферентні.
Кальцифіли — рослини, вибагливі до вмісту кальцію в середовищі. Серед судинних макрофітів до кальцифілів належить елодея канадська.
Кальцифоби — рослини, що уникають багатого карбонатного живлення. До них належать сфагнові мохи, водяний горіх плаваючий (Тгара паі;ащ) і ін.
Індиферентні — рослини, байдужі до наявності кальцію в середовищі (грунті).
Мікроелементи необхідні рослинам в дуже малих кількостях, але мають величезне значення для їх життя, оскільки входять до складу багатьох життєво важливих ферментів. Причому при невеликій потребі рослин в мікроелементах їх вміст в навколишньому середовищі також малий. Мікроелементи нерідко виступають як лімітуючі чинники. До них відносяться 10 елементів: Fe, Мп, Гп, Си, В, Si, Мо, СІ, V, Со (залізо, марганець, цинк, мідь, бор, кремній, молібден, хлор, ванадій і кобальт).
Такі елементи, як мідь, марганець, цинк, фтор, що зустрічаються в водоймах у малих кількостях, відіграють значну роль у житті водних організмів (вплив на ріст, дихання, обмін, харчування, розмноження та ін.) При збільшенні вмісту цих речовин у воді до певних концентрацій, вони стають токсичними для гідробіонтів.
В організмі рослин мідь входить до складу ферментів-оксидаз та білка пластоціаніну. Надлишок сполук міді має токсичну дію. Занурені рослини накопичують важкі метали (Мп, Си, Сг) в 4-9 разів більше, ніж повітряно-водні.
Іони заліза та марганцю зустрічаються в природних водах у вигляді гідрокарбонатів, сульфатів, хлоридів, фосфатів, а залізо також у гуміновому комплексі. Нестача заліза у ґрунті або порушення його засвоєння затримують розвиток рослин, призводять до виникнення хлорозів та інших патологічних аномалій. Несприятливо впливає на рослини і надлишок заліза.
Водорості різних відділів мають неоднакові потреби в макро- і мікроелементах. Так, для нормального розвитку діатомових водоростей необхідні досить значні кількості кремнію, який використовується для побудови їх панцирю. За відсутності або нестачі кремнію панцирі діатомових стоншуються.
11.3. Екологічне значення біотичних факторів
Між живими організмами, що населяють атмосферу, літосферу і гідросферу, складаються відносини, при яких кожний з компонентів впливає на інші безпосередньо або внаслідок зміни екологічного середовища і виступає в ролі біотичного фактора. Тому всю сукупність різноманітних і опосередкованих впливів рослин, тварин і мікроорганізмів один на одного називають біотичними факторами. Вони надзвичайно різноманітні і включають фітогенні, зоогенні і мікробогенні дії, серед яких виникають прямі й опосередковані, антагоністичні і симбіотичні відносини, механічні й хімічні дії.
Фітогенні фактори - впливи інших рослин-співмешканців: прямий (механічні контакти, симбіоз, паразитизм, поселення епіфітів), так і опосередкований (фітогенні зміни середовища існування рослин - виявляються у формі перехоплення води, поживних речовин грунту, вуглекислого газу та світла, необхідних для фотосинтезу тощо).
Зоогенні фактори - вплив тварин (поїдання, витоптування і інші механічні впливи, запилення квітів, розповсюдження насіння, опосередкований вплив на середовище).
Мікогенні фактори - вплив грибів (паразитів і симбіонтів).
Водний фітоценоз відзначається специфічним видовим складом, який включає вищі й нижчі рослини. Між ними створюються складні фітоценотичні взаємовідносини, які викликаються фітогенни-ми факторами.
Серед представників рослинного царства є види, що належать до факультативних або облігатних гетеротрофів, які одержують готову органічну речовину. До них належать сапрофіти, паразити та комахоїдні рослини.
Сапрофітний спосіб живлення передбачає використання органічної речовини відмерлих залишків рослин і тварин. Цей спосіб живлення широко відомий у водоростей, але трапляється й у покритонасінних.
Паразитизм рослин — одна з форм співжиття різних організмів,
з яких один, втративши здатність до синтезу органічних речовин, живе за рахунок іншого. Паразитизм буває постійний і тимчасовий. Паразити поселяються на поверхні або в середині тіла рослини і живляться її соками. Вони позбавлені справжньої кореневої системи або ж вона дуже редукована, і замість неї утворюються кореневі присоски, за допомогою яких паразити поглинають воду, пластичні речовини, вітаміни. Серед паразитів є бактерії, гриби і квіткові рослини.
Флора покритонасінних включає чимало паразитних і напівпара-зитних видів. До цієї групи належать види повитиці, вовчкових і петрів хрест. Вони паразитують на трав'яних і чагарникових рослинах, а деякі на деревах.
Напівпаразитами називаються рослини, що поселяються на інших рослинах, за рахунок яких частково живляться, частково самостійно виробляють поживні речовини (льонолисник, ремнецвітник європейський, омела біла і ін.).
Рослини істотно впливають на тварин. Відомо понад 500 видів рослин, які живляться за рахунок тварин, зокрема комах. Вони створюють специфічну групу комахоїдних рослин. Це зелені рослини, які мають спеціальні пристосування для ловлі й перетравлювання різних дрібних тварин, найчастіше комах. Так вони доповнюють своє автотрофне живлення однією з форм гетеротрофного живлення. Найчастіше такі рослини трапляються на грунтах, де замало азоту, тому готова органічна речовина для них насамперед є джерелом азотного живлення. Такі рослини приваблюють комах певним забарвленням, запахом, солодкими виділеннями, захоплюють їх специфічними ловильними механізмами пасивного або активного типу, а потім виділяють спеціальні ферменти, які забезпечують внутрішньоклітинне перетравлення.
В Україні трапляються комахоїдні рослини з родини росичкових - росички (Drosera rotundifolia, D. longifolia). У росички є речовини, які характеризуються паралітичною дією на комах (рис. 11.5). Після прилипання бічні волоски росички нахиляються до жертви і виділення посилюється.
Листки росички мають хлорофіл і здійснюють фотосинтез. Якщо ж цю рослину позбавити тваринної їжі, вона хоча і не гине, проте розвивається дуже повільно. Процес всмоктування продуктів розкладу забезпечується видільними залозами, що з'єднані з провідною системою рослини.
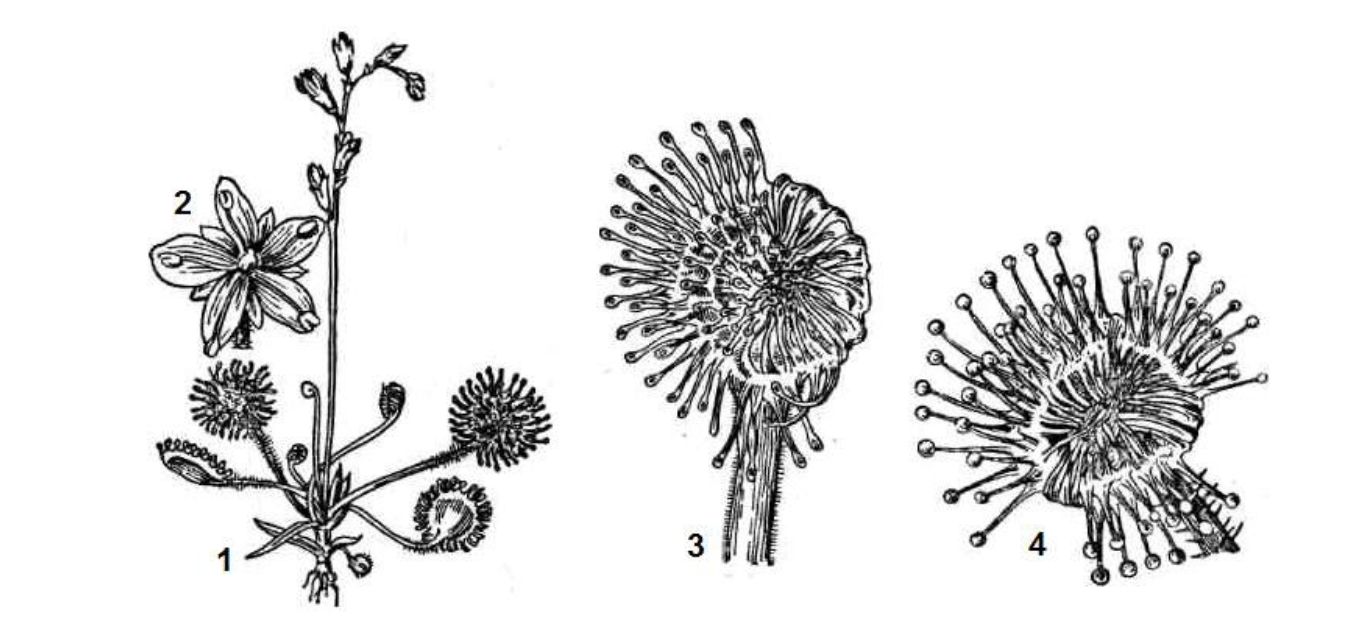
Рис. 11.5. Росичка круглолиста (Drosera rotundifolia):
1 - загальний вигляд, 2 - квітка, 3 - окремий листок, 4 - половина війок заломлена до центру, де знаходиться комаха
Специфічні ловильні апарати мають пухирники (Utricularia). Це розгалужені прикореневі листки, що мають глечикоподібну форму (рис. 11.6). Вздовж країв вони мають щетинки, які заважають комасі вийти з ловильного апарату. Комаха, що потрапила в ловильний апарат, гине і розчиняється під дією ферментів, що виділяються листком, а її соки поглинаються всмоктуючими клітинами, які вистилають стінки пастки.
Відомо більш як 400 видів покритонасінних рослин, здатних одержувати поживні речовини подібним способом.
Симбіоз рослин — це форма співіснування різних організмів, від якого обидва компоненти одержують взаємну користь. Симбіоз дуже поширений у природі і буває облігатний і факультативний.
Облігатний симбіоз виявляється між бобовими рослинами і ві-льноіснуючими азотфіксуючими. Бульбочкові бактерії зв'язують вільний азот повітря в солі азотної кислоти, які доступні для споживання вищими рослинами. Самі ж бактерії в процесі життєдіяльності одержують від бобових готові органічні речовини. Ці симбіотичні відношення між рослинними організмами людина використовує в практичній діяльності. Посіви бобових збагачують грунт на азот, потім ці площі використовуються для сівби злакових, які позбавлені цієї можливості.

Рис. 11.6. Пухирник (иігіеиіагіа)
1 - загальний вигляд; на підводному стеблі розвиваються у великій кількості дрібні бульбашки;
2 - бульбашка в збільшеному вигляді; 3 - бульбашка в розрізі — видно клапан (а), вії (б) і всмоктуючі клітини (в);
4 - всмоктуючі клітини при великому збільшенні
Мікориза — це співжиття, асоціація кореня вищих рослин і непатогенного гриба. Мікотрофний тип живлення характерний для рослин, які мають мікоризу на коренях і добувають поживні речовини із ґрунту за допомогою гіфів грибів, симбіотично зв'язаних з кореневою системою.
За таких взаємовідносин обидва партнери приносять користь один одному. До симбіозу з грибами здатні більшість деревних і багато трав'янистих рослин. Як правило, гриби інфікують молоді корені. При цьому кореневі волоски в них не розвиваються. Гриб одержує від вищої рослини органічні речовини і своєю чергою забезпечує доступність для рослин води та мінеральних речовин. Гриб допомагає вищій рослині у засвоєнні важкодоступних речовин гумусу. Своєю ферментативною системою гриб сприяє обміну вуглеводів, активізує діяльність ферментів вищих рослин. Явище мікоризи дуже поширене у природі.
Особливий інтерес представляють випадки співжиття водоростей з іншими організмами. Найчастіше водорості використовують живі організми як субстрат. За характером субстрату, на якому поселяються водорості обростань, серед них виділяють епіфіти, що поселяються на рослинах, і епізоїти, що живуть на тваринах. Так, на вапнякових черепашках молюсків можна зустріти види родів Сіайорйога або Оейоропіиш, в обростаннях губок звичайні деякі зелені, синьозелені і діатомові водорості. В угрупованнях обростань між рослиною-господарем і рослиною, що утворює обростання, встановлюються неміцні і короткочасні зв'язки.
Водорості можуть жити також в тканинах інших організмів — за межами клітини (у слизі, міжклітинниках водоростей, іноді в оболонках мертвих клітин), і внутріклітинно. Водорості, що живуть в тканинах або в клітинах інших організмів, називають ендофітами. Позаклітинні і внутріклітинні ендофіти з числа водоростей утворюють досить складні симбіози — ендосимбіози. Для них характерна наявність постійних і міцних зв'язків між партнерами. Ендосим-біонтами можуть бути різні водорості — синьозелені, зелені, бурі, червоні та інші, але найчастіше зустрічаються ендосимбіози одноклітинних зелених і жовтозелених водоростей з одноклітинними тваринами. Водорості, що беруть участь в них, мають назву зоохлорел і зооксантел.
Жовтозелені і зелені водорості утворюють ендосимбіози і з багатоклітинними організмами — прісноводними губками, гідрами і ін. Своєрідні ендосимбіози синьозелених водоростей з найпростішими і деякими іншими організмами одержали назву синціанозів. При цьому виникає морфологічний комплекс, який називають ціаномом, а синьозелені водорості в ньому — ціанелами.
Вищий ступінь розвитку симбіотичних відносин характеризується строгою постійністю компонентів симбіотичної системи. Наприклад, у тілі прісноводної папороті азолли (Л/о11а 61іси1оійе8), незалежно від того, де б вона не розвивалася, завжди поселяються колонії певного виду — АпаЬаепа агоііае. Всі спроби зараження азолли представниками інших родів або видів синьозелених водоростей успіху не мали. Це свідчить про те, що між учасниками даного симбіозу існує специфічний фізіологічний взаємозв'язок.
Ендосимбіотичний спосіб життя, як правило, призводить до зміни зовнішнього вигляду учасників симбіозу. Так, у синьозеленої
водорості Aphanocapsa, що живе в тканинах морської губки Aplisilla, товщина оболонки значно менша, ніж у вільноживучих видів синьозелених водоростей. Ще серйознішим змін зазнають водорості — внутріклітинні ендосимбіонти: вони втрачають оболонку, будова їх джгутиків спрощується, зникає стигма, так що часто стає неможливим встановити таксономічну належність водорості.
Серед симбіозів, що утворюються водоростями, найбільший інтерес представляє їх симбіоз з грибами, в результаті якого виникла своєрідна група рослинних організмів - лишайники. Цей симбіоз демонструє унікальну біологічну єдність, яка призвела до появи принципово нового організму. Разом з тим кожен партнер симбіозу лишайника зберігає риси тієї групи організмів, до якої він відноситься. Лишайники є єдиним доведеним випадком виникнення нового організму в результаті симбіозу двох.
Конкуренція. На розвитку окремих видів водоростей можуть позначатися і відносини конкуренції. Так, бурі водорості з роду фукус зазвичай мешкають в зоні припливів і зазнають періодичного (іноді до двох діб) пересихання. Нижче, в постійно затоплюваній зоні, як правило, розташовуються щільні зарості інших бурих і червоних водоростей. Проте в тих місцях, де ці зарості не дуже щільні, фукуси виростають і на більшій глибині.
Алелопатія — це хімічний вплив речовин одних видів рослин на інші за допомогою фізіологічних виділень. Рослини виділяють леткі або краплинно-рідкі речовини, при цьому одні види стимулюють або гальмують розвиток інших.
Бентосні водорості, наприклад, починають чинити взаємний вплив з моменту осідання і проростання спор. Експериментально доведено, що зооспори ламінарії (Laminaria) не проростають поблизу з фрагментами слані бурої водорості з роду аскофілюм (Ascophylum).
Мікроорганізми виділяють маразміни — фізіологічно активні речовини, що зумовлюють в'янення і старіння рослин. Разом з тим мікроорганізми і гриби виділяють антибіотики, гіберелін і дуже отруйні алкалоїди. Бактерії, що продукують антибіотики, при високій концентрації їх можуть отруюватися, а при незначній — стимулювати свій розвиток.
Отже, рослинні організми через стимулюючі і гальмівні виділення здатні регулювати чисельність і балансувати взаємозв'язки між ценобіонтами в угрупованнях.
Зоогенні фактори. Значний вплив на життя рослин мають тварини. Відносини між ними складаються насамперед на базі живлення.
У більшості випадків водорості в екосистемах виступають як продуценти органічної речовини. У зв'язку з цим найважливішим чинником, що обмежує розвиток водоростей в конкретній екосистемі, є наявність консументів, що існують за рахунок поїдання водоростей. Наприклад, розвиток угруповань з домінуванням видів роду Laminaria біля Атлантичного узбережжя Канади лімітується чисельністю морських їжаків, що харчуються переважно цими водоростями. У тропічних водах в зонах коралових рифів зустрічаються райони, в яких риби повністю виїдають зелені, бурі і червоні водорості з м'якою сланню, залишаючи нез'їденими синьозелені водорості з жорсткими оболонками. Черевоногі молюски також в основному харчуються водоростями. Повзаючи по дну вони поїдають мікроскопічні водорості і проростки макроскопічних видів. При масовому розвитку цих молюсків можуть відбуватися серйозні порушення у водоростевих угрупованнях літоралі.
До фітофільних риб відноситься білий амур, який споживає водні рослини (переважно вищі), і тому його називають «трав'яним коропом». Білий амур акліматизований в Україні і використовується у полікультурі риб.
Винятково важливі біоценотичні відносини виникають між ентомофільними рослинами і тваринами-запилювачами. При запиленні комахи живляться нектаром або пилком. Щоб одержати нектар або пилок їм потрібно проникнути в квітку. У цієї рослин-ентомо-філів квітка побудована так, що під час збору нектару комахи вимазують тіло пилком і запилюють ним приймочки маточки інших квіток, які відвідують. Так відбувається перехресне запилення.
11.4. Антропогенні чинники
Як і всяка інша жива істота, людина як ланка біоценозу є біотичним чинником для решти організмів екосистеми, в якій вона знаходиться.
Вплив людини на природу може бути свідомим і стихійним (випадковим). Людина свідомо виводить нові сорти рослин і породи тварин, знищує «шкідливі» види, створює нові біоценози. Проте рівень знань людства про функціонування екосистем не завжди дозволяв зробити правильний прогноз наслідків антропогенного втручання у природу. Як правило, у історії людства свідомий антропогенний вплив на природу приносить позитивні плоди на перших етапах. Пізніше проявляються негативні наслідки для екосистем: хижацьке винищення окремих промислових організмів; непродумане переселення видів, які, конкуруючи з аборигенними, змінюють екосистеми; розорювання земель, що руйнує природні біоценози і знижує видову різноманітність; забруднення водних екосистем продуктами хімізації сільського господарства тощо.
До випадкових належать впливи, які раніше не були заплановані і передбачені: найнебезпечніші екологічні катастрофи - аварії в вугільній, нафто- і газовидобувній галузях, на підприємствах металургійної, хімічної, нафтохімічної і мікробіологічної промисловості. До числа найбільших екологічних катастроф минулого століття відноситься аварія на Чорнобильській АЕС 1986 року, яка, крім безпосередніх наслідків (загибель персоналу і ліквідаторів, радіаційне забруднення території), зумовила випадіння радіоактивних опадів, які різко збільшили ризик онкозахворювань у населення Європи.
Серйозними факторами випадкового руйнівного впливу є природні катастрофи трансформованого людиною середовища (землетруси, суховії, смерчі, повені).
Менші за розміром антропогенні впливи - випадкове завезення тварин і насіння бур’янів у інші природні зони і території.
Особливо шкідливими є техногенні і урбогенні процеси.
Прокладаючи канали і споруджуючи водосховища, людина створює нові місця для водних організмів, що нерідко принципово відрізняються від водойм даного регіону за гідрологічним і тепловим режимом. На даний час рівень продуктивності багатьох континентальних водойм часто визначається не стільки природними умовами, скільки суспільними і економічними відносинами. Скидання стічних вод нерідко приводять до збіднення видового складу і загибелі водних рослин або до масового розвитку окремих видів. Перше відбувається при скиданні у водойму токсичних речовин, друге — при збагаченні водойми біогенними речовинами (особливо сполуками азоту і фосфору) в мінеральній або органічній формі — т.з. антропогенне евтрофування водойм. У багатьох випадках стихійне збагачення водойм біогенними речовинами відбувається в такому
масштабі, що водойма як екологічна система виявляється переобтяженою ними. Наслідком цього є надмірний бурхливий розвиток водоростей — «цвітіння» води.
На водорості, особливо аерофітні і грунтові, можуть мати вплив і атмосферні викиди токсичних промислових відходів. Часто наслідки мимовільного або цілеспрямованого втручання людини в життя екосистем мають необоротний характер.
Разом із сполуками азоту, фосфати сприяють антропогенному евтрофуванню водойм. Але, через невисоку рухомість у ґрунті, фосфорні сполуки є менш небезпечними для навколишнього середовища.
11.5. Життєві форми рослин
Життєва форма (екоморфа, біоморфа) - це зовнішній вигляд і біологічні особливості рослин, які відбивають їх пристосованість до певних умов середовища існування.
Існують різні класифікації життєвих форм рослин, найпопулрні-шою з яких є класифікація за К. Раункієром (1906-1907 рр.). Ця система побудована на ознаці, що характеризує пристосування рослин до перенесення несприятливої пори року (холодної або сухої) за положенням бруньок відновлення на рослині по відношенню до рівня грунту і снігового покриву.
К. Раункієр виділив п’ять головних життєвих форм: фанерофіти, хамефіти, гемікриптофіти, криптофіти і терофіти.
У фанерофітіе бруньки зимують або переносять сухий період «відкрито», і досить високо над землею (дерева, кущі, дерев’янисті ліани, епіфіти), тому переважно захищені спеціальними лусочками, що зберігають конус наростання і молоді зачатки листків від втрати вологи.
У хамефітів бруньки розташовуються майже на рівні грунту або не вище 20-30 см над ним (кущики, півкущики, рослини, що стеляться). У холодній і помірній кліматичних зонах ці бруньки зимують під снігом.
У гемікриптофітів бруньки відновлення перебувають на рівні грунту, або занурені у грунт чи підстилку з опалого листя та решток рослин на деяку глибину. Це трав’янисті багаторічні рослини.
Криптофіти - представлені або геофітами з бруньками відновлення в землі на деякій глибині (кореневища, бульби і цибулини), або гідрофітами з бруньками відновлення, що зимують під водою.
Криптофіти - це геофіти водні.
У терофітів усі вегетативні частини відмирають до кінця сезону і зимуючих бруньок не лишається. Вони відновлюються на наступний рік з насіння.
К. Раункієр з’ясував взаємозв’язок життєвих форм з кліматом. У вологих тропіках переважають фанерофіти, у помірній зоні - гемікриптофіти, у пустелях - терофіти, у тундрі і пустелях - хамефіти.
У ході еволюції рослинного світу змінювалися і життєві форми. Перші наземні рослини, що вийшли з води на сушу були невисокими рослинами, подібними до сучасних трав’янистих. Пізніше розвивалися великі деревовидні форми, у тому числі папороті, 30-45 м висотою. Були і трав’янисті папороті, що дожили до наших днів. Голонасінні представляють собою деревну групу. Квіткові рослини - найрізноманітніші за життєвими формами. У ході еволюції покритонасінні пройшли шлях від порівняно невисоких з товстими стовбурами, малорозгалужених розеточних дерев (подібних до сучасних пальм і динного дерева) до великих справжніх дерев з добре розвинутим стовбуром і кроною; від дерев - до кущів, кущиків і різноманітних трав. Трав’янисті рослини більш пристосовані до освоєння нових екологічних умов.
11.6. Стратегії рослин
Є різні способи виживання популяцій рослин в угрупованнях і екосистемах, для характеристики яких використовується термін «стратегії рослин». За типом стратегій рослин в угрупованнях Ра-менський Л.Г. у 1938 році виділив три типи: віоленти, патієнти та експлеренти. Віоленти - це види «леви», зростають у сприятливих і не порушених місцях, домінанти і едифікатори рослинних угруповань з широкими реалізованими нішами, об’єм яких наближається до ніш фундаментальних. Це потужні рослини з розвинутою кореневою системою, що дозволяє їм ставити під контроль ресурси еда-фічного середовища і світла. Віоленти не мають спеціальних пристосувань для переживання несприятливих умов і реагують на погіршення умов зменшенням розмірів, негативно реагують на порушення, оскільки не мають банків насіння або банків вегетативних зачатків. Патієнти - види «верблюди», рослини несприятливих місцезростань, мають здатність до перенесення несприятливих умов за рахунок спеціальних фізіолого-біохімічних механізмів пережи
вання стресу. Експлеренти - види-волоцюги, або «шакали», мають високу насіннєву продуктивність, здебільшого посилену інтенсивним вегетативним розмноженням. Як правило, це рослини з укороченим життєвим циклом. Експлеренти володіють слабкою конкурентною здатністю, мешканці порушених місцезростань, формують найперші стадії автогенних сукцесій.
У 1979 році Дж. Грайм, не знаючи про попередню роботу Рамен-ського, заново описує ці ж три типи стратегій під назвою конкурентів (К-статегів), стрес-толерантів ^-статегів) і рудералів ^-статегів). Дж. Грайм виділив три типи стратегій і назвав їх первинними, вказавши, що існують вторинні типи стратегій, і через це поділ рослин досить умовний. На даний час типи стратегій Раменсь-кого-Грайма - це один з найпопулярніших варіантів системи стратегій рослин.
Існує система стратегій рослин, яка базується на співвідношенні витрат енергії на репродуктивне зусилля і підтримання вегетативного стану рослин - типи стратегій Мак-Ліода-Піанки. Мак-Ліод був першим ученим, який у 1884 році підійшов до питання про існування у рослин типів стратегій і виділив категорії «пролетаріїв» і «капіталістів». Англійський дослідник Е.Піанка у 1970 р. практично перевідкрив типи стратегій Мак-Ліода, розробивши концепцію двох полярних типів К і г, які виділяються відносно частки життєвої енергії, яка використовується на розмноження. У першому випадку (К), основна життєва енергія рослини йде на підтримання вегетативної активності (на ріст і підтримання організму в дорослому стані), а в другому (г) - велика кількість енергії використовується на розмноження, на «виробництво» великої кількості потомства, яке викидається навмання, і вид виживає завдяки потужному «банку насіння».
Найпоширеніші в Україні вищі водні рослини можна віднести до таких типів:
К-стратеги (віоленти) - очерет звичайний, рогіз широколистий, глечики жовті, рдесники блискучий і плаваючий;
8-стратеги (патієнти) - ряска мала, латаття біле, комиш озерний;
К-стратеги (експлеренти) - рдесник кучерявий, спіродела бага-токоренева, частуха подорожникова.
Багато видів перехідні між двома або навіть трьома стратегічними типами. Наприклад, кушир занурений і водопериця колосиста -Б-И-стратеги, а рдесник пронизанолистий потрапляє в змішану групу KRS.
11.7. Рослини - індикатори змін природного середовища
Комплекс екологічних факторів середовища існування досить строго визначає, які організми можуть жити в даному місці, а які не можуть. Використавши обернену закономірність, можна робити висновки про середовище існування за організмами, які в ньому проживають. Науковий напрямок оцінки екологічних факторів середовища існування за допомогою живих організмів, їх популяцій і угруповань називають біоіндикацією. Ті живі організми, життєві функції яких так тісно корелюють з певними факторами середовища, що можуть застосовуватися для їх оцінки, називають біоіндикаторами. Розширене поняття біоіндикатора включає не лише власне біологічні системи, а і їх структури (популяції видів, угруповання) і ознаки, функція яких корелює з певними факторами середовища.
Методи біоіндикації широко використовують для оцінки антропогенного впливу на природу. Про сам екологічний фактор за результатами біоіндикації можна судити лише опосередковано. При оцінці забруднення навколишнього середовища вона доповнює традиційні фізико-хімічні дослідження аналізом біологічних наслідків дії антропогенних чинників.
У зв’язку з біологічними особливостями рослин, фітоіндикацію виділяють в окрему велику галузь біоіндикації. За допомогою рослин і їх угруповань оцінюють різні фактори антропогенного впливу: порушення природних ландшафтів, забруднення повітря газоподібними речовинами (лишайники, окремі види хвойних), забруднення водного середовища промислово-комунальними стоками тощо. Є істотні методичні відмінності використання у ролі фітоіндикаторів вищих і нижчих рослин різного систематичного положення.
Якість або ступінь забруднення води за складом водних рослин оцінюють двома способами: 1) за індикаторними організмами; 2) за результатами порівняння структури угруповань на ділянках з різним ступенем забруднення і контрольному. У першому випадку за присутністю або відсутністю індикаторних видів або груп і їх відносної кількості, користуючись наперед розробленими системами індикаторних організмів, відносять водойму або її ділянку до певного класу вод. У другому випадку висновок роблять за наслідками порівняння складу водних рослин на різних ділянках водойми, в різній мірі схильних до забруднення.
Наприклад, у водних фітоценозах р. Устя, яка протікає через м. Рівне і у яку потрапляють комунально-побутові стічні води обласного центру, на різних ділянках змінюється число видів занурених рослин (від 10-12 на ділянках з відносно чистою водою, до 1-2 на забруднених). Із виявлених на річці видів найчутливішими до забруднення виявилися водопериця колосиста, латаття біле, ряска триборозенчаста і рдесник блискучий. За їх наявністю або відсутністю можна зробити висновок про якість води річки. Загалом, найчутливішими до забруднення води вважаються занурені рослини, у яких контакт з водним середовищем максимальний, а найстійкіши-ми - гелофіти, прибережні повітряно-водні рослини
Ступінь забруднення водних об'єктів органічними речовинами визначає їх сапробність (8арго8 - гниючий), а розділ гідроекології, що вивчає такі забруднення, називається сапробіологія.
Видова структура угруповань гідробіонтів залежно від їх чутливості до органічного забруднення водойм чітко виражена на біоце-нотичному рівні. Тому в 1908 р. німецькі дослідники К. Кольквітц і Р. Марссон вважали, що розклад органічної речовини, що знаходиться у складі стічних вод, має ступінчастий характер. У зв'язку з цим водні об’єкти або їх зони залежно від ступеня забруднення органічними речовинами підрозділяють на полі, - мезо- і олігосапробні.
У полісапробній зоні, що знаходиться поблизу від місця скидання стічних вод, відбувається розщеплення білків і вуглеводів в анаеробних умовах. Ця зона характеризується майже повною відсутністю вільного кисню, наявністю у воді білків, що не розклалися, значних кількостей сірководню і діоксиду вуглецю, відновним характером біохімічних процесів. Число видів водоростей, здатних розвиватися в цій зоні, порівняно невелике, але вони зустрічаються в масових кількостях.
У мезосапробній зоні забруднення виражене слабше: білків, що не розклалися, немає, сірководню і діоксиду вуглецю небагато, кисень присутній в помітних кількостях, проте у воді ще є аміак, аміно-і амідокислоти. Мезосапробна зона підрозділяється на а- і Р-мезосапробні підзони. У першій зустрічаються аміак, аміно- і амідокислоти, але вже є кисень. У цій зоні зустрічаються синьозелені водорості родів осциляторія і формідіум. Мінералізація органічної ре-
човини в основному йде за рахунок аеробного окиснення, зокрема бактерійного. Наступна мезосапробна підзона характеризується присутністю аміаку і продуктів його окиснення — азотної і азотистої кислот. Амінокислот немає, сірководень зустрічається в незначних кількостях, кисню у воді багато, мінералізація йде за рахунок повного окиснення органічної речовини. Видова різноманітність водоростей тут вища, ніж у попередній підзоні, але чисельність і біомаса організмів нижчі. Найбільш характерними для цієї підзони є діатомові водорості з родів мелозіра, діатома, навікула і зелені з родів космарі-ум, спірогіра, кладофора, сценедесмус.
У олігосапробній зоні сірководень відсутній, діоксиду вуглецю мало, кількість кисню наближається до нормального насичення, розчинених органічних речовин практично немає. Для цієї зони характерна висока видова різноманітність водоростей, але чисельність і біомаса їх незначні.
За висновком чеського ученого Сладечека В., який у 1973 році склав список вищих водних рослин у системі сапробності, судинні макрофіти розвиваються переважно в олігосапробній та Р-мезосапробній зонах.
Використання водоростей як індикаторних організмів у геології і юридичній практиці базується на наявності серед них великої кількості стенотопних видів, що потребують для існування певної комбінації зовнішніх умов, характерних для певного біотопу, поза яким вони не можуть існувати. На противагу їм широко розповсюджені евритопні види, здатні існувати в різних біотопах, як індикаторні організми непридатні. Використання водоростей як індикаторних організмів в геології, крім того, можливе завдяки їх великій старовині, а також здатності деяких з них зберігатися у викопному стані.
11.8. Вищі водні рослини у декоративній аквакультурі
Значення вищих водних рослин у декоративній аквакультурі часто виявляється не менш вагомим, ніж риб і інших водних тварин. Це пов’язано із тим, що поряд із включенням макрофітів у процеси трансформації органічних сполук, насиченням води киснем та ін., вищі водні рослини у аквакультурі відіграють ще й неоціненну декоративну роль. Саме завдяки рослинам, що є елементами декору, вдається надати будь-якому акваріуму естетичного та привабливого вигляду. Для окремих видів риб рослини у нерестовому акваріумі
служать нерестовим субстратом або матеріалом для побудови гнізда, окремі об’єкти декоративної аквакультури (молодь риб, дрібні риби, ракоподібні) знаходять сховище у заростях рослин.
Значення декоративної складової рослин у акваріумі підтверджується популярними напрямками акваріумістики, у яких основний акцент робиться на створення довершеної композиції із водних рослин, моделювання певних природних біотопів окремих груп макрофітів. У таких напрямках інші об’єкти декоративної аквакультури стають другорядними та розглядаються як додаткові складові біотопу акваріумів.
У декоративній аквакультурі поширені переважно тропічні рослини. Саме ці повітряно-водні та водні рослини краще підходять для вирощування та розведення, оскільки більшість із них характеризуються стабільним ростом протягом усього року, відсутністю періодів спокою. Асортимент декоративних водних рослин постійно розширюється завдяки ввезенню і акліматизації нових видів та селекційній роботі акваріумістів. Серед таких рослин є досить невибагливі у культивуванні, які не вимагають значних зусиль для вирощування та розведення, є й надскладні для культивування у декоративній аквакультурі. Основними факторами середовища, на які необхідно зважати при вирощуванні рослин у декоративній аквакультурі, є освітлення, фізико-хімічні властивості води та видовий склад гідробіонтів. Таким чином, декоративні рослини мають певні вимоги до середовища утримання: яскрава чи помірна освітленість, температура в межах 20-30°С, якість води, наближена до якості природної водойми, де росте даний вид, та відсутність активних шкідників - рослиноїдних риб і інших тварин, що пошкоджують або висмикують рослини. Основними критеріями підбору рослин для декоративного акваріуму, відповідно, є можливість адаптації рослини до умов штучної водойми - акваріуму, привабливий вигляд та розміри, співставні із розміром самого акваріуму.
Створення акваріумного пейзажу із вищих рослин вимагає знання їх біології, особливостей росту і розмноження, вимог до основних умов утримання. Компонування рослин може здійснюватись на основі особистих уподобань акваріуміста, видового складу певного природного біотопу, який моделюється у акваріумі, але підбір повинен здійснюватися із врахуванням можливості спільного утримання у обмеженому об’ємі акваріума за визначених параметрів
середовища. Так, спільне культивування у одному акваріумі рослин, що потребують відносно невисокої температури води (20-25°С) та порівняно теплолюбних рослин, що краще розвиваються при температурі від 26 до 30°С можливе лише при умові забезпечення певного компромісу. Найкращим рішенням є підбір видів, що мають схожі вимоги до температури. Ще важливішими параметрами водного середовища для водних рослин є значення рН та загальна твердість води. Спільне утримання видів рослин, які у природних водоймах пристосовані до діаметрально протилежних параметрів, є досить проблематичним. Водночас у декоративній аквакультурі можна ефективно утримувати рослини, що потребують різної інтенсивності освітлення або швидкості течії. У такому випадку рослини, що потребують затінку висаджують на слабко освітлених ділянках акваріуму (бічні та задня стінки) або у затінку потужних та високих рослин, що потребують яскравого освітлення. Оскільки у акваріумі можна створити ділянки із порівняно потужною циркуляцією води завдяки використанню повітродувного або фільтруючого обладнання, при компонуванні рослин, які різняться за вимогами до руху води, можна вчинити аналогічно.
Водні рослини, які вирощуються у декоративній аквакультурі, поділяють на такі групи: плаваючі на поверхні води (не укорінені, з надводним або із підводним листям), короткостеблові (переважно укорінені, водні або повітряно-водні) та довгостеблові (переважно не укорінені, повітряно-водні, рідше - водні).
До справжніх водних рослин, які здатні розвиватись лише у водному середовищі, належить відносно невелика группа видів. Найпоширенішими є представники родин жабурникових (роди егерія -Egeria, елодея - Elodea, гідрила - HydriПa, жабурникові - Hydro-charis, лагаросифон - Lagarosiphon, лімнобіум - Limnobium, блікса -Blyxa, оттєлія - Ottelia та валлінерія - VaШsneria), лататтєвих, апо-ногетонових, рдесникових, куширових, столисникових (водопериці - MyriophyПum) і амарилісових (крінуми - Crinum).
Більшу частину асортименту акваріумних рослин складають болотні (повітряно-водні) рослини, які у природних умовах ростуть і у воді, і над водою (різні екофази). Залежно від сезонних коливань рівня води рослини у процесі адаптації формують листкову пластинку іншої форми, ніж у підводній екофазі. Темпи росту рослин у повітряному середовищі зазвичай значно перевищують швидкість
росту у воді, тому на ринку декоративної аквакультури переважають рослини, вирощені у вологих оранжереях. Водночас асортимент декоративних рослин постійно поповнюється новими видами із природних водойм тропіків та субтропіків. Серед швидкоросту-чих рослин цієї групи можна виділити представників родин амарантових (альтернатери - Alternathera), шорстколистих (бакопи - Ba-copa, хеміантус - Hemianthus), акантових (гігрофіли - Hygrophila), онагрових (людвигії - Ludwigia), лобелієвих (лобелії - Lobelia), плакунових (ротали - Rotala, аманії - Ammannia, незеї - Nesaea). Ці рослини відносяться до довгостеблових неукорінених.
Короткостеблові болотні рослини також характеризуються достатньо високими темпами росту у порівнянні із справжніми водними макрофітами. Найпоширенішими у акваріумах є представники родів ехінодорус (Echinodorus), криптокорина (Cryptocoryne), анубіас (Anubias); також водні папороті та мохи, що займають відокремлену нішу в акваріумах.
Серед окремих ареалів природного поширення декоративних рослин можна виділити найяскравіші, або характерні для певної систематичної групи рослин. Саме такі біотопи найчастіше моделюють у декоративних акваріумах - моделях прибережних ділянок природних водойм. Розглянемо деякі з них.
1. Річка Ріо-Гвапоре (Бразилія). Навколо неї ще й досі існують ліси, не змінені негативним впливом цивілізації. Ґрунт на багатьох прибережних ділянках складається із дрібного білого піску, найбільше різноманіття рослин спостерігається на замулених ділянках, із повільною течією. Уздовж берега поширені великі види ехінодорусів (Echinodorus paniculatus, E. grandiflorus), а також низькорослі види, що краще підходять для культивування у акваріумі, але потребують досить яскравого освітлення (Echinodorus tenellus, E. bolivianus). Також на ділянках, що періодично затоплюються, біля узбережжя поширені повітряно-водні людвигії (Ludwigia sedoides). Із справжніх водних рослин найхарактернішими для прозорого мілководдя даного біотопу є кабомби (Cabomba furkata), лімнофіли (Limnophila indica), та ейхорнії (Eichornia diversifolia, E. azurea, E. crassipes). На порівняно глибоких ділянках (до 2 м) зустрічається отелія (Ottelia brasiliensis). Плаваючі рослини переважно представлені лімнобіумом та сальвінією.
2. Заплавні райони р. Ріо-Сипао (Венесуела). Води даної річки
характеризуються високою каламутністю. Серед ґрунтів переважають важки суглинки із високим вмістом заліза. Найпоширеніші на прибережних ділянках бакопи, тоніна та ейхорнія (Eichornia diversi-folia). Як і в інших водоймах Південної Америки, тут поширені ехінодоруси, окремі види яких зростають на періодично затоплюваних ділянках, інші - у напівзануреному стані, значно менша кількість видів - у зануреному стані.
3. Річки басейну Ріо-Уругвай (Аргентина). Найбільша різноманітність та чисельність спостерігається на ділянках із глинистими грунтами і глибиною до 2 м. На узбережжях при слабкій течії домінують численні види кабомб, водопериць, людвигій. Для ділянок із швидкою течією характерними є гігрофіли та Eichornia azurea. Порівняно глибокі ділянки заселяють німфоїди. Порівняно прозора вода таких річок та водотоків дозволяє розвиватись елеоха-рисам, водоперицям та егерії.
4. Річки та стоячі водойми Центральної Африки. Головною прикрасою декоративних акваріумів, що походить із даних водойм, є рослини із роду анубіас (Anubias). Більшість видів є болотними рослинами, що ростуть на поверхні або у напівзануреному стані, окремі здатні розвиватись під водою, не виходячи на сушу. Умови освітлення на різних ділянках дуже різняться, тому більшість анубіасів добре пристосовані до затінку, водночас можуть добре рости і при яскравому освітленні. У водоймах та прибережній смузі зустрічаються також папороті, аманії та крінуми.
5. Східноафриканські озера тектонічного походження: Малаві і Таньганьїка. Біотопи із водними рослинами знаходяться у зоні тростини та на літоралі (ділянки із найменшими коливаннями води). Для цих водойм на ділянках глибиною 0,5-4 м характерними є валіснерія спіральна, кушир занурений, водопериця, рдесники. Порівняно рідко у водоймах даного типу можна зустріти гідрилу (Hydrilla verticillata) та наяди (Najas horrida, N. marina).
6. Біотоп річок острову Мадагаскар. Піщані грунти із значними включеннями глини обумовлюють характерне глинисто-мутне забарвлення води у річках та інших водоймах цього острова. Найяскравішими представниками водної флори острова є апоногетони. Види, які ростують у тимчасових водоймах, що часто пересихають; або ті, що розвиваються у водоймах зі стрімкою течією, досить рідко культивують у акваріумах через наявність у рослин чітко виражених періодів спокою. Епізодично такі рослини потрапляють із природних біотопів у мережі зоомагазинів. Центральною рослиною штучно створеного біотопу (акваріуму) такого типу є апоногетон мадагаскарський (Aponogeton madagascariensis), який є однією із найиопулярніших у акваріумістиці рослин.
7. Біотоп водотоків Південної Азії. Невеликі водойми даного регіону, вода яких переважно має склабкокислу реакцію та невисокий вміст солей кальцію та магнію, є природним ареалом існування представників роду криптокорина (Сгурїосогупе) - популярних та широко розповсюджених у акваріумах рослин. Як і більшість повітряно-водних рослин тропіків, криптокорини зустрічаються у зануреному стані, на поверхні біля водойми або на затоплених ділянках водойм. У таких місцях переважають глинисті грунти із високим вмістом заліза, окремі ділянки являють собою мулисті наноси. Фізико-хімічний склад води у таких водотоках протягом року змінюється незначно, що обумовлює певні біологічні особливості криптокорин: в умовах декоративної аквакультури у разі несприятливих умов або значному коливанні параметрів води можна спостерігати досить неприємне явище - «криптокоринову хворобу». У водоймах Південної Азії зростають також гігрофіли, папороті, мохи та лататтєві.
Останнім часом розведення та вирощування вищих водних рослин у штучних умовах (оранжереї, басейни) для потреб декоративної аквакультури все більше переважає над їх збиранням у місцях природного поширення. Таким чином, можна стверджувати, що сучасна акваріумістика, окрім основного завдання, має перспективу сприяти збереженню видового різноманіття природних ділянок водойм, що піддаються антропогенному впливу. Досягненням декоративної аквакультури можна вважати також виведення нових видів та сортів рослин, які не зустрічаються у природі, але вже давно завоювали популярність в акваріумах.
11.9. Основи фітоценології
Поняття про фітоценоз. Сукупність організмів, взаємозв'язаних і взаємообумовлених загальним обміном речовин і енергії, називають біоценозом. Рослини, що населяють біоценоз, створюють фітоценоз, тварини — зооценоз, а мікроорганізми — мікробоценоз.
Фітоценоз, рослинне угруповання — сукупність рослинних організмів на відносно однорідній ділянці, які перебувають у взаємодії між собою, з тваринами і навколишнім середовищем. Часто трапляються поєднання, що виникають внаслідок співжиття багатьох видів вищих і нижчих рослин. Фітоценоз є основною одиницею рослинності. Кожний фітоценоз характеризується певним видовим складом. Морфологічна структура фітоценозу визначається просторовим розподілом рослин.
Фітоценоз є динамічною системою, якій властива сезонна і різ-норічна мінливість компонентів під впливом екологічних умов.
Наука, що вивчає рослинні угруповання (фітоценози), - фітоценологія. Це розділ геоботаніки і біогеоценології (багато ботаніків ототожнюють фітоценологію з геоботанікою).
Сформоване рослинне угруповання характеризується певним флористичним складом, структурою, ценотичними взаємовідношеннями і екологічною пластичністю його компонентів. У зв'язку з цим фітоценоз відрізняється також певною ярусністю, покриттям, рясністю, життєвістю, фізіономічністю, періодичністю, сталістю виду тощо.
Компоненти фітоценозу відрізняються за рівнем їх участі в фітоценозі, переважно за створенням фітомаси і проективним вкриттям. Домінантами називаються переважаючі види, які завдяки оптимальним умовам інтенсивно розвиваються і займають провідне місце в угрупованні як за кількістю особин, так і за фітомасою. Кількість їх різна і залежить від структури ценозів, його флористичного складу і віку. У вертикальному розрізі водних рослинних угруповань розрізняють домінантів надводного, плаваючого і підводного ярусів. Такі рослинні угруповання називають полідомінантними. Угруповання, де окремі види складають 80-90%, називаються монодомінантними, а при переважанні двох видів — бідомінантними.
В ценозах прибережно-водних рослин, як правило, домінують рогози вузьколистий, широколистий та очерет звичайний і визначають загальний аспект водотоку.
Види рослин, що відіграють важливу роль в складі угруповання, але менш істотну, ніж домінанти, цього ярусу, називають співдомі-нантами. Наприклад, у непроточних або малопроточних ділянках водойм Полісся України в угрупованнях дрібних вільноплаваючих рослин домінантою є ряска мала, а співдомінантою — спіродела багатокоренева.
За впливом на фітоценотичне середовище розрізняють едифікатори - це види, які контролюють режим відносин угруповання і визначають його (можуть домінувати в угрупованні, але не обов’язково), і види-супутники (асектатори), які мало впливають на створення середовища всередині угруповання. Наприклад, осока є едифікатором в осоково-різнотравних угрупованнях. Асектатором прибережних повітряно-водних фітоценозів може бути паслін солодко-гіркий або незабудка болотна.
Водні фітоценози переважно складаються невеликою кількістю видів, особливо у водоймах з низькою прозорістю води. Часто вони складаються лише з одного виду або з невеликою участю інших видів. Це чисті або майже чисті одноярусні фітоценози цього виду (зарості очерету звичайного або рогозу вузьколистого).
Маловидові спрощені угруповання досить широко представлені серед усіх груп водних рослин, а у водоймах з низькою прозорістю води вони переважають над іншими, складніше влаштованими угрупованнями. У водоймах з високою прозорістю води, крім одно-видових одноярусних ценозів, зустрічаються ценози з одним або декількома домінуючими видами, до яких приєднується велика кількість супутніх видів, які розчленовані на декілька ярусів.
Синузія — екологічно і просторово відокремлена частина рослинного угруповання (фітоценозу). Складається з рослин однієї або кількох екологічно близьких життєвих форм, іноді синузію може становити одна популяція.
Вертикальна структура фітоценозів. Вертикальне розчленування рослинного угруповання за рахунок контрастних за висотою життєвих формам рослин називають ярусністю, а екологічно відособлені структурні частини - ярусами.
Ярусність — характерна ознака рослинного угруповання. За несприятливих умов існування розвиваються одноярусні угруповання, а за оптимальних — багатоярусні.
У вертикальному розчленуванні водних рослинних угруповань розрізняються три основних яруси:
- надводний з під'ярусами (або ярусами) за висотою: високих, середніх і низьких надводних рослин;
- плаваючий з вільноплаваючих і укорінених рослин з плаваючим листям;
- підводний з під'ярусами (або ярусами) за висотою: високих, се
редніх і маленьких придонних рослин (трав).
Розподіл рослин за ярусами визначається їх вибагливістю до екологічних факторів. В одному ярусі ростуть види, близькі за вибагливістю до умов освітленості, мінерального живлення тощо, тобто види екологічно рівноцінні.
Ярусність рослинного угруповання має велике біологічне значення. Завдяки їй на обмеженій ділянці поселяються і співіснують види різної екологічної вибагливості, виникають стійкі фітоценози, що вирізняються високою продуктивністю.
Рясність. Характерною рисою рослинного угруповання є рясність виду — кількісна участь його особин у формуванні ценозу. Вона залежить від біологічних властивостей виду, умов місцезростання і фітоценотичних якостей і може бути виражена кількістю особин на одиницю площі, масою органічної речовини, площею, розселення та ін.
Рясність виду залежить від інтенсивності його життєвих процесів, здатності розвивати фотосинтезуючу і поглинальну поверхні. Завдяки цьому вони легко конкурують з іншими рослинами за світло воду, мінеральні речовини і досягають значного розвитку.
Рясності виду сприяє насінне і вегетативне розмноження. У флорі водойм, як правило, переважають багаторічні рослини з потужними кореневищами, здатні до інтенсивного вегетативного розмноження, які витісняють інші рослини. Наприклад, елодея канадська, яку за швидке вегетативне розмноження назвали «водяною чумою».
Фітоценотичні властивості видів також обумовлюють рясність видів в угрупованні, що чудово ілюструється взаємовідношенням між видами різних ярусів. Високі і густі зарості прибережно-водних рослин істотно впливають на розвиток занурених рослин шляхом світлової конкуренції, енергії відновлення, життєвості виду тощо. Вищі водні рослини в результаті насичення води фізіологічними виділеннями та іншими конкурентними властивостями, пригнічують розвиток водоростей.
Покриття. Відображає геометричну структуру фітоценозу. Воно є характерною ознакою фітоценозу, відображає боротьбу рослин за простір і раціональне використання світла, вологи, тепла, мінерального живлення. Види, що досягають верхніх ярусів і розвивають величезну поверхню повітряного живлення, синтезують більше органічної речовини та енергії, ніж ті, які ростуть під ними. Вони до
сягають високої рясності та істотної ролі в структурі угруповання і кругообігу речовин. Неоднорідність покриття пов'язана з ярусним розподілом видів угруповання і сезонними змінами клімату.
Покриття — це просторово виражена величина горизонтальної проекції надземних органів рослин на зайняту ними поверхню землі. Розрізняють проективне і справжнє покриття. Проективне покриття - це частка покриття виду від загального покриття угруповання. Найчастіше покриття виражають у відсотках. Справжнє (істинне) покриття створене основами стебел рослин (наприклад після викошування травостою). Справжнє покриття буде завжди меншим від проективного покриття.
Життєвість (віталітет) - рівень життєвого стану рослини, який забезпечує реалізацію генетично обумовленої програми росту і розвитку. Це одна з характерних ознак, що свідчить про екологічне і фітоценотичне пристосування його компонентів до життя в рослинному угрупованні. Для позначення життєвості виду часто користуються такою 5-бальною шкалою:
5 - вид у рослинному угрупованні знаходить оптимальні умови, має пишний розвиток і підвищене плодоношення та цвітіння;
4 — цей вид має все для розвитку і завершує цикл плодоношенням та активним висівом насіння;
3 — вид має хороший вегетативний розвиток, але не проходить всього життєвого циклу (не досягає звичайних своїх розмірів і не висіває насіння);
2 — вид спочатку нормально розвивається, добре вегетує, але не плодоносить;
1 — вид з самого початку незадовільно вегетує, не плодоносить і дуже пригнічений.
Життєвість виду в фітоценозі обумовлюється його конкурентною здатністю, стійкістю проти хвороб і шкідників, здатністю до симбіозу з іншими організмами, пристосованістю до мінливих умов повітряно-ґрунтового і водно-мінерального живлення.
Періодичність розвитку рослин у фітоценозі виявляється у зміні його аспектів.
Аспект - зовнішній вигляд фітоценозу, його фізіономічність в певний період його розвитку.
Аспект визначається зовнішністю всіх видів, що населяють угруповання. Нерідко визначається участю виду і станом розвитку
його вегетативних і генеративних органів. Фізіономічність багатьох рослинних угруповань озер і водосховищ визначається вегетацією і цвітінням лататтєвих (глечиків жовтих і латаття білого), плодоношенням рогозових тощо.
Фізіономічність має певну динамічність - у помірній зоні змінюється кілька разів протягом вегетації (сезонний аспект).
Хронологічна аспектність зумовлена не тільки біологічними властивостями (періодом плодоношення, цвітіння), а й екологічними умовами. Так, поява і цвітіння чи плодоношення окремих видів на піщаних обмілинах р. Дніпра визначаються часом їх звільнення з-під води і настанням оптимальних грунтово-повітряних умов.
Сталість виду. Ця характерна ознака рослинності визначається ступенем поширення виду на різних ділянках даного фітоценозу.
Так, в угрупованні занурених рослин із рдесника пронизанолис-того останній вид зустрічається на всіх 20 вивчених ділянках даного угруповання, рдесник гребінчастий — на 10, кушир занурений — на 7, а водопериця колосиста — на 2 ділянках. Отже, сталість цих видів у фітоценозі неоднозначна і, виражаючи її в %, у рдесника про-низанолистого вона складає 100%, рдесника гребінчастого — 50, куширу — 35, водопериці — 10%.
Види рослин, що зустрічаються на більшості (понад 50%) досліджених ділянок даного фітоценозу, називають константними. Види, що зустрічаються на 25% досліджених ділянок фітоценозу, називаються другорядними, а види з сталістю менше 25% — випадковими. При детальному вивченні рослинності кількість досліджених ділянок повинна бути не менше 20 одиниць для кожного фітоценозу.
Розміщення виду. У фітоценозі види розміщуються по-різному. Більшість особин одного виду більш-менш рівномірно розподіляється на всій площі, другого — скупчені плями, третього — групами або плямами. Ця ознака виду зумовлена його біологічними особливостями та характером умов місцезростання.
Контрольні запитання та завдання
1. Як впливають на рослинний організм умови його місцезростання?
2. Як класифікують екологічні фактори і яка їх роль у розвитку рослин?
3. Які є екологічні групи рослин відносно світла, води, температури?
4. Які пристосування виникають у рослин до водного середовища існування?
5. Як впливає на рослини хімічний склад води?
6. Яке екологічне значення біотичних факторів для рослин?
7. Як впливає на водні рослини людина?
8. Які водорості індикують різні зони сапробності?
9. Що називають стратегіями рослин?
10. Які типи стратегій водних рослин виділяють?
11. Яке значення водних рослин у декоративній аквакультурі?
12. Які види рослин найчастіше використовують для оформлення декоративних акваріумів?
13. Що називають фітоценозом? Назвіть ознаки фітоценозу.
14. Що таке флористичний склад і структура фітоценозу?
15. Що називають життєвістю виду?
16. Які види називають домінантами, співдомінантами, едифікаторами і асектаторами?
17. Поясніть значення термінів: синузія, рясність, покриття, ярусність, аспект.
