СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 14. ОДНОКЛЕТОЧНЫЕ ПРОТИСТЫ: ВОДНЫЕ ПЛЕСЕНИ, СЛИЗЕВИКИ, ХИТРИДИОМИЦЕТЫ И ОДНОКЛЕТОЧНЫЕ ВОДОРОСЛИ
Характеристики отделов
В данной главе будут рассмотрены четыре отдела гетеротрофов и три отдела автотрофов. Гетеротрофы чрезвычайно разнообразны. Среди них хитридиомицеты и оомицеты имеют ценоцитные (многоядерные, но не разделенные перегородками) гифы, которые у некоторых оомицетов могут быть длинными и сильно разветвленными. Оомицеты образуют подвижные споры бесполого размножения (зооспоры) с двумя жгутиками — гладким и перистым (см. рис. 2-26), тогда как зооспоры хитридиомицетов имеют один гладкий задний жгутик. Миксомицеты, пока не размножаются, большую часть жизни проводят в виде текучего скопления цитоплазмы. Акразиомицеты — это амебовидные организмы, не образующие жгутиковых клеток ни на одной стадии жизненного цикла. Характерные черты этих гетеротрофных отделов и автотрофных протистов приводятся в табл. 14-1.
Последние также довольно сильно различаются по строению жгутиков (если они есть) и биохимическим особенностям, в частности по составу пигментов, запасным веществам и компонентам клеточных стенок. Эти признаки (в том числе и для организмов, рассмотренных в гл. 15) представлены в табл. 14-1. Названия некоторым отделам водорослей даны по окраске преобладающих вспомогательных пигментов, которые маскируют зеленый цвет хлорофиллов. При фотосинтезе вспомогательные пигменты отдают энергию поглощенного ими света главным образом в фотосистему II. Среди этих организмов обнаружено большое разнообразие запасных продуктов; у большинства это различные углеводы, часто вместе с липидами. На гетеротрофное происхождение автотрофных протистов указывает то, что некоторые роды хризофитов, динофлагеллят и эвгленовых регулярно захватывают твердые частицы нищи, как и представители многих отделов, которые не приобрели в ходе эволюции симбиотических хлоропластов.
Таблица 14-1. Сравнительная характеристика 10 отделов протистов
Отдел |
Число видов |
Фотосинтстические пигменты |
Запасные углеводы |
Жгутики |
Компоненты клеточной стенки |
Среда обитания |
Oomycota (водные плесени) |
475 |
Нет |
Гликоген |
2; гладкий и перистый; Целлюлоза только у репродуктивных клеток |
Водные или нуждаются в воде |
|
Chytridiomycota (хитридиомицеты) |
750 |
Нет |
Гликоген |
1 задний гладкий; только у репродуктивных клеток |
Хитин, другие полимеры |
Водные |
Acrasiomycota (клеточные слизевики) |
70 |
Нет |
Гликоген |
Нет (амебоидное движение) |
Нет |
Наземные |
Myxomycota (плазмодиальные слизевики) |
450 |
Нет |
Гликоген |
2 гладких; только у репродуктивных клеток |
Нет |
Наземные |
Chrysophyta (хризофиты) |
6650 |
Хлорофиллы а и с, каротиноиды, включая фуко- ксантин |
Хризоламина рин |
0, 1 или 2; апикальные, гладкие или перистые, равные или неравные |
Нет или целлюлоза, у некоторых с кремне- земовыми чешуйками; кремнезем у диатомей |
Морские и пресноводные |
Pyrrhophyta (динофла- гелляты) |
1100 |
Хлорофиллы а и с, каротиноиды, включая периди- нии |
Крахмал |
0 или 2 боковых перистых |
Целлюлоза, другие вещества |
Морские и пресноводные |
Phaeophyta (бурые водоросли)1 |
1500 |
Хлорофиллы а и с, каротиноиды, включая фукок- сантин |
Ламинарии, маннитол |
2 боковых, перистый направлен вперед, гладкий назад; только у репродуктивных клеток |
Целлюлозный матрикс с альгиновыми кислотами (полисахариды) |
Почти все морские ; очень обильны в холодных океанах |
Rhodophyta (красные водоросли)1 |
4000 |
Хлорофилл а, каротиноиды, фико- билины |
Крахмал |
Нет |
Целлюлоза, пектиновые вещества, у многих карбонат кальция |
Морские, некоторые пресноводные, много тропических видов |
Chlorophyta (зеленые водоросли)1 |
7000 |
Хлорофиллы а и b,каротиноиды |
Крахмал |
0, 2 или больше; верхушечные или боковые, равные, гладкие |
Полисахариды, иногда целлюлоза |
В основном пресноводные, некоторые морские |
Euglenophyta (эвгленовые |
800 |
Хлорофиллы а и b,каротиноиды |
Парамилон |
От 1 до 3, верхушечные, перистые (с одним рядом боковых волосков) |
Нет клеточной стенки; имеют белковую пелликулу |
Большинство пресноводные |
1 Отделы, состоящие в основном или целиком из многоклеточных организмов; описаны в гл. 15.
Различные группы протистов накапливают в результате фотосинтеза весьма неодинаковые углеводы и липиды. У зеленых водорослей в хлоропластах накапливается крахмал (как и у их потомков — растений); у всех прочих групп запасные вещества откладываются в цитоплазме. У красных водорослей и динофлагеллят так запасается крахмал, у бурых водорослей — полиглюкозид ламинарии, имеющий другие связи между остатками глюкозы. В других отделах с хлорофиллом с основной запасной продукт — хризоламинарин, более полимеризованная форма ламинарина. Многие бурые водоросли накапливают также маннитол, спиртовое производное сахара маннозы. Эвгленовые синтезируют парамилон (полисахарид спиральной конфигурации) и этим резко отличаются от других отделов.
Отдел Oomycota
Отдел оомицетов с примерно 475 видами — весьма специфическая группа протистов. Клеточные стенки этих организмов образованы в основном целлюлозой или целлюлозоподобными полимерами, что резко отличает их от грибов. Хромосомы оомицетов напоминают хромосомы большинства эукариот и совсем не похожи на высококонденсированные хромосомы грибов. Мейоз и митоз аналогичны этим процессам у других эукариот; присутствуют центриоли. Этот отдел охватывает от одноклеточных до сильно разветвленных ценоцитных нитчатых форм.
Многие виды оомицетов могут размножаться как половым, так и бесполым путем. Половое размножение, оогамия, включает образование оогония, содержащего множество яйцеклеток, и антеридия с многочисленными мужскими ядрами (рис. 14-4). Сингамия приводит к образованию зиготы с толстой стенкой (ооспоры), которая является покоящейся спорой; эта структура дала название всему отделу. Как уже упоминалось, бесполое размножение у оомицетов происходит посредством подвижных зооспор с двумя жгутиками— одним гладким, другим перистым.
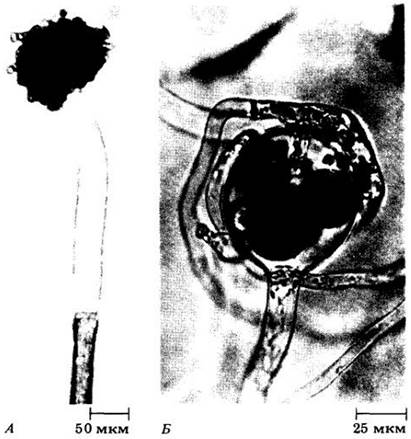
Рис. 14-4. Асhlyа ambisexualis. водная плесень с половым и бесполым размножением. А. Пустой спорангий с зооспорами, образовавшими цисты у его отверстия (отличительная черта Асhlyа). Б. Половые органы. Показано, как оплодотворяющие трубочки антеридия проникают через стенку оогония к яйцеклеткам
Крупная группа оомицетов — водные организмы. Ее представители, называемые водными плесенями, в изобилии обитают в пресной воде и легко могут быть выделены из нес. Многие из них сапробионты, но известны и паразиты, в том числе вызывающие болезни рыб и ее икры.
У некоторых водных плесеней (например, у Saprolegnia, рис. 14-5) половой процесс происходит путем образования мужских и женских половых органов на одной и той же особи, т. е. эти организмы гомоталломны. Другие виды (например, в роде АсЫуа, рис. 14-4) гетероталломны, т. с. мужские и женские половые органы развиваются на разных экземплярах, а если на одном, то генетически процесс самооплодотворения невозможен. Как Saprolegnia, так и Achlya могут размножаться бесполым и половым путями.
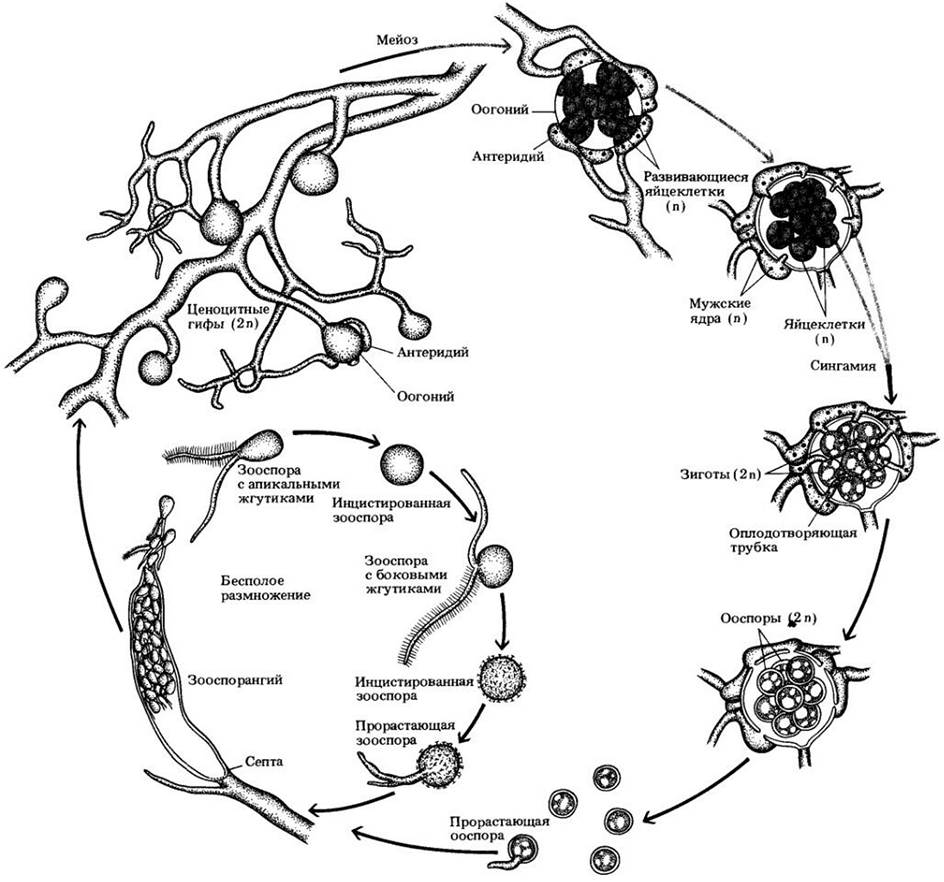
Рис. 14-5. Жизненный цикл оомицета Saprolegnia. Мицелий этой водной плесени диплоидный. Размножение в основном бесполое. Двужгутиковые зооспоры выходят из зооспорангия, некоторое время плавают, а затем превращаются в цисту. Каждая из них в конечном итоге дает начало вторичной зооспоре, которая также инцистируется, а затем прорастает, давая начало новому мицелию. При половом размножении оогонии и антеридии образуются на одной и той же гифе. Внутри них происходит мейоз. Оогонии — это разросшиеся клетки, в которых образуется несколько сферических яйцеклеток. Антеридии развиваются из верхушек других нитей той же особи и содержат многочисленные мужские ядра. При созревании антеридии растут по направлению к оогониям и образуют проникающие в них оплодотворяющие трубочки. Мужские ядра движутся по этим трубочкам к женским ядрам и сливаются с ними, в результате чего возникает толстостенная зигота — ооспора. Прорастая, она развивается в гифу, которая затем образует зооспорангий, и цикл начинается снова
Другая группа оомицетов является в основном сухопутной, хотя организмы в присутствии капельножидкой воды также образуют подвижные зооспоры. Сюда относится порядок Peronosporales, несколько представителей которого имеют важное экономическое значение. Как сказал С. Дж. Алексопулос: «По крайней мере два из них приложили руку, или, точнее говоря, гифы к формированию хозяйственной истории существенной части человечества».
Вид Plasmopara vitícola вызывает заболевание, называемое ложной мучнистой росой винограда, или мильдью. Эта болезнь была случайно занесена во Францию в конце 1870-х гг. на американской виноградной лозе, которая импортировалась из-за устойчивости к другим болезням, и вскоре поставила под угрозу все виноделие этой страны. Борьба с мильдью, наконец, увенчалась успехом благодаря счастливому случаю и внимательным наблюдениям. Крестьяне в окрестностях Медока обычно опрыскивали свои виноградные посадки вдоль дорог смесью медного купороса и извести с неприятным вкусом, чтобы гроздья не обрывали прохожие. Профессор из университета в Бордо, изучавший проблему мильдью, заметил, что такие растения не заболевают. После совещания с владельцами виноградников он приготовил особую смесь химикатов, названную бордосской жидкостью, которая стала широко применяться с 1882 г. Это первый химический препарат, использованный для борьбы с болезнями растений.
Другой экономически важный представитель оомицетов — род Phytophthora (что по-гречески означает «разрушитель растений»), включающий 35 видов и являющийся особенно злостным фитопато геном, поскольку в широких масштабах поражает многие сельскохозяйственные культуры, включая какао, ананасы, томаты, каучуконосы, папайю, лук, землянику, яблони, сою, табак и цитрусовые. Хорошо известный представитель этого рода — Phytophthora cinna- momi, обитающий в почве, несколько лет назад уничтожил или сделал неурожайными миллионы авокадовых деревьев в южной Калифорнии и других местах. Он также погубил десятки тысяч гектаров ценных посадок эвкалиптов в Австралии. Зооспоры Р. cinnamomi привлекаются к поражаемым растениям химическими выделениями их корней. Этот оомицет образует устойчивые споры, сохраняющиеся до шести лет во влажной почве. Сейчас ведутся интенсивные селекционные работы по созданию устойчивых к нему сортов авокадо и других культур.
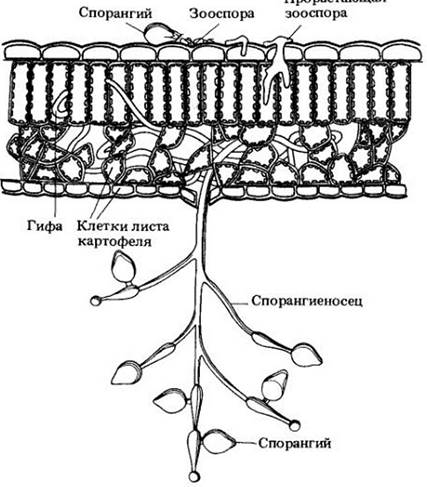
Однако больше всего «прославился» вид Phytophthora infestans (рис. 14-6) — возбудитель фитофтороза картофеля, ставший причиной тяжелого голода в Ирландии в 1846 — 1847 гг. Население страны, увеличившееся между 1800 и 1845 гг. с 4,5 до 8,5 млн. человек, сократилось в результате этого бедствия до 6,5 млн. в 1851 г. Около 800 тыс. жителей погибло от истощения, остальные эмигрировали, в основном в США. Практически все посадки картофеля в Ирландии погибли летом 1846 г. в течение одной недели, что явилось катастрофой для местных крестьян, питавшихся почти исключительно картофелем из расчета 4 — 6 кг в день на взрослого человека (такой рацион был необходим для получения достаточного для нормальной жизнедеятельности количества белка).
Рис. 14-6. Phytophthora infestans, возбудитель фитофтороза картофеля. Клетки листа показаны серым цветом. В присутствии воды при низкой температуре зооспоры выходят из спорангиев и плывут до места прорастания (как показано здесь) или же сами спорангии прорастают ростковой трубкой
Другой представитель этого отдела — «голубая плесень» (Peronospora hyoscyami) — причинил в 1979 г. табачным плантациям США и Канады убыток примерно в четверть миллиарда долларов. Этот вид распространяется многоядерными спорами, легко переносимыми по воздуху. В будущем можно предполагать возрастание числа подобных эпидемий, так как генетическая изменчивость многих культур сужается. Эта проблема будет обсуждаться в гл. 30.
Отдел Chytridiomycota
Хитридиомицеты, включающие около 750 видов, обитают главным образом в воде. Эти организмы чрезвычайно разнообразны по форме, природе своих половых процессов, а также жизненным циклам. Их клеточные стенки образованы в основном хитином, хотя в них встречаются и другие полимеры. Митоз и мейоз, насколько известно, аналогичны протекающим у оомицетов. Все хитридиомицеты являются ценоцитными, но при созревании у них появляется несколько септ.
Основная характерная черта хитридиомицстов — единственный гладкий задний жгутик их подвижных клеток (зооспор и гамет). Эта особенность позволяет выделять их среди прочих протистов и указывает на родство между различными формами, объединяемыми в данный отдел.
Некоторые хитридиомицеты — простые одноклеточные организмы, не образующие мицелия. У них все тело в подходящее время становится репродуктивной структурой. Другие имеют тонкие ризоиды для прикрепления к субстрату (рис. 14-7). Различные виды хитридиомицетов паразитируют на водорослях, водных оомицетах, спорах, пыльцевых зернах и других частях сосудистых растений; другие, будучи сапробионтами, обитают, например, на мертвых насекомых.
Рис. 14-7. Chyiridium confervae, широко распространенный хитридиомицет (снимок сделан с помощью оптической системы Номарски). Заметны тонкие, распространяющиеся вниз ризоиды
Род Соelomomyces объединяет облигатных паразитов личинок москитов и других двукрылых. Его жизненный цикл напоминает цикл ржавчинных грибов, так как происходит со сменой хозяев — мелких водных рачков из отряда веслоногих и личинок насекомых. Coelomomyces сейчас изучается как возможное средство биологической борьбы с москитами.
Строение и размножение других хитридиомицетов гораздо сложнее. Рассмотрим, например, жизненный цикл рода Allomyces. У некоторых его видов происходит чередование изоморфных поколений (см. рис. 14-8), у других — гетероморфных, когда гаплоидные и диплоидные особи неодинаковы. Чередование поколений характерно для растений и многих водорослей, но среди прочих живых существ обнаружено только в роде Allomyces, в другом близко родственном ему роде хитридиомицетов (см. рис. 10-10), а также у очень большого числа гетеротрофных протистов, не рассматриваемых в этой книге. С точки зрения морфологии, физиологии, цикла развития, а также половых гормонов Allomyces является одним из наиболее изученных протистов.
Рис. 14-8. У хитридиомицета Allomyces arbusculus, чей жизненный цикл представлен на рисунке, происходит чередование изоморфных поколений. Гаплоидные и диплоидные особи неразличимы до начала формирования органов размножения. На гаплоидных образуется примерно равное число бесцветных женских и оранжевых мужских гаметангиев. Мужские гаметы привлекаются гормоном сиренином, выделяемым в два раза более крупными женскими гаметами. Зигота теряет жгутики и прорастает, давая диплоидный организм. Этот спорофит образует два типа спорангиев: (1) бесполые — бесцветные тонкостенные структуры, высвобождающие диплоидные зооспоры, которые в свою очередь прорастают снова в диплоидное поколение; (2) половые — толстостенные красновато-бурые структуры, способные выдержать суровые условия среды. После периода покоя в них происходит мейоз, и возникают гаплоидные зооспоры. Эти зооспоры развиваются в гаплоидные особи, которые, созревая, образуют гаметангии
Отдел Acrasiomycota
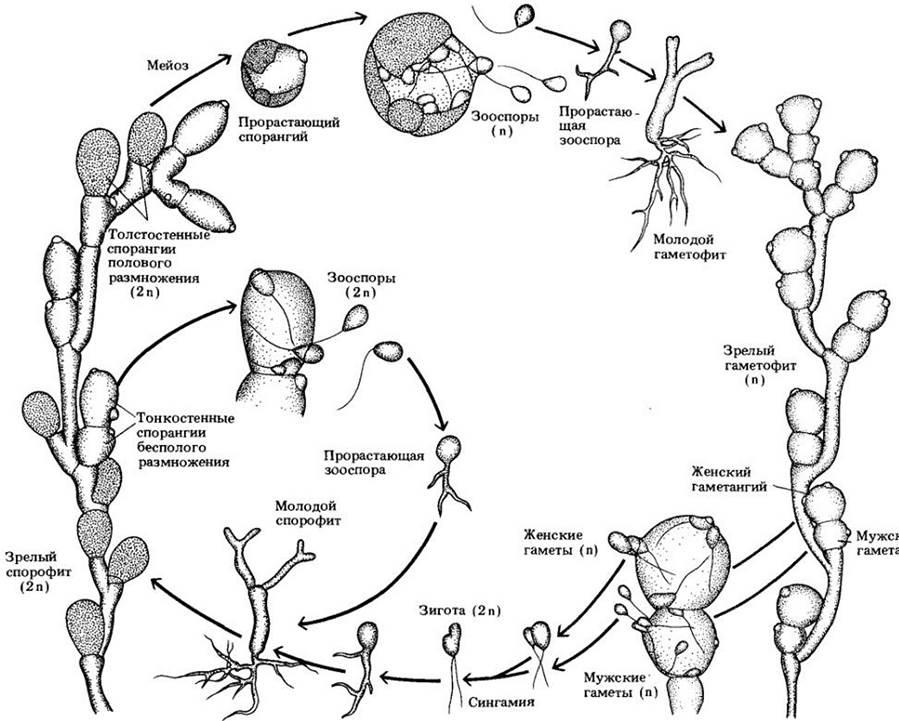
Клеточные слизевики — группа из 65 видов, объединяемых в несколько родов, — возможно, ближе к корненожкам (тип Rhizopoda), чем к другим организмам. В противоположность грибам их клеточная стенка богата целлюлозой, а митоз сопровождается исчезновением ядерной оболочки; имеются центриоли. Один из представителей этого отдела Dictyostelium discoideum обсуждался в гл. 8 в качестве модели клеточной дифференцировки. У этого и других видов акразиомицетов амебы (миксамебы) сохраняют свою индивидуальность, даже агрегируясь на определенной стадии жизненного цикла с образованием «слизистого» псевдоплазмодия (рис. 14-9). Миксамебы могут также по отдельности инцистироваться (формировать микроцисты) и в такой форме переживать краткие периоды засухи или голода.
Рис. 14-9. Dictiostelium discoideum. А. Агрегация миксамеб. Заметно, что каждая амеба сохраняет свою индивидуальность. Стрелка указывает направление, в котором движется поток миксамеб. Б. Общий вид большого количества агрегировавших амеб при малом увеличении
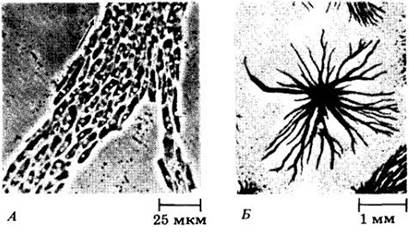
Бесполое размножение спорами — обычное явление среди клеточных слизевиков. Часто наблюдается и половое размножение с участием макроцист. У Dictyostelium discoideum,например, макроцисты являются несколько уплощенными, неправильно округлыми до эллиптических, многоклеточными структурами диаметром 25 — 50 мкм (рис. 14-10). В процессе их образования пары гаплоидных амеб сначала сливаются, давая зиготы, которые затем привлекают и захватывают амеб, находящихся поблизости. Возникающие агрегаты меньше, чем в случае формирования «слизей», и имеют скорее округлые, а не продолговатые очертания. Каждая макроциста окружается тонкой мембраной, выделяемой амебами. Позднее вся группа клеток внутри мембраны покрывается толстой клеточной стенкой, богатой целлюлозой. В макроцисте зигота (единственная в жизненном цикле диплоидная клетка) проходит перед прорастанием мейоз и несколько митотических делений, в результате чего образуются и выходят наружу новые гаплоидные амебы.
Рис. 14-10. Процесс образования макроцисты у Dictyostelium mucoroides. А. Каждая зигота, или гигантская клетка, начинает поглощать находящиеся вокруг амебы. Б. Гигантские клетки поглотили всех амеб и покрылись целлюлозной стенкой. В. Зрелые макроцисты; на этой стадии их содержимое кажется однородным
Клеточные слизевики считались раньше редкостью. В 1933 г. К. Рейпер открыл Dictyostelium discoideum; его дальнейшие исследования этого организма привлекли большое внимание к этой группе. Сейчас известно, что она включает обычных обитателей большинства почв и особенно лесной подстилки, где они питаются бактериями. Как и большая часть различных типов организмов, акразиомицеты лучше представлены в тропиках, чем в умеренной зоне. Клеточных слизевиков легко выделять и культивировать. Они оказались прекрасным экспериментальным материалом для изучения клеточной дифференцировки и молекулярной биологии (см. с. 124 — 125).
В 1982 г. Д. Уодделл из Принстонского университета описал необычное свойство разводимого в культуре хищного акразиомицета Dictyostelium caveatum, которого он выделил из экскрементов летучих мышей в пещере Бланчард-Спринге в шт. Арканзас. Амебы этого вида образовывали общие агрегаты с амебами всех испытанных видов клеточных слизевиков, подавляли их развитие и поедали, формируя в конечном итоге плодовые тела, состоящие только из клеток своего вида.
Отдел Мyxomycota
Этот отдел объединяет плазмодиальных слизевиков, или миксомицетов, — всего около 450 видов, — по-видимому, не имеющих прямых связей ни с клеточными слизевиками, ни с другими группами протистов. В подходящих условиях миксомицеты существуют в виде тонкой текучей массы протоплазмы, передвигающейся как амеба. Лишенная клеточной стенки, эта «голая» протоплазма называется плазмодием. Плазмодии, перемещаясь, поглощают и переваривают бактерий, дрожжевые клетки, споры грибов и мелкие частицы разлагающихся растений и животных. Успешное культивирование плазмодиев в среде без оформленных частиц пищи позволяет предполагать, что они могут питаться и путем абсорбции.
Плазмодий может достигать массы 20 — 30 г, но поскольку распространяется чрезвычайно тонким слоем, то это количество может покрыть поверхность в несколько квадратных метров (см. рис. 10-5, А). Он содержит много ядер, не разделенных клеточными стенками. По мере роста происходит многократное синхронное их деление. Имеются центриоли, и митоз обычный, но хромосомы очень мелкие.
Как правило, движущийся плазмодий напоминает по форме веер с текучими протоплазматическими трубочками, которые толще у его основания, а к наружному краю ветвятся и утончаются. Эти трубочки образованы слегка затвердевшей протоплазмой, сквозь которую быстро перетекает ее более жидкая фракция. Передний край плазмодия представлен тончайшей пленкой геля, отделенного от субстрата только плазматической мембраной и слизистой оболочкой неизвестного химического состава.
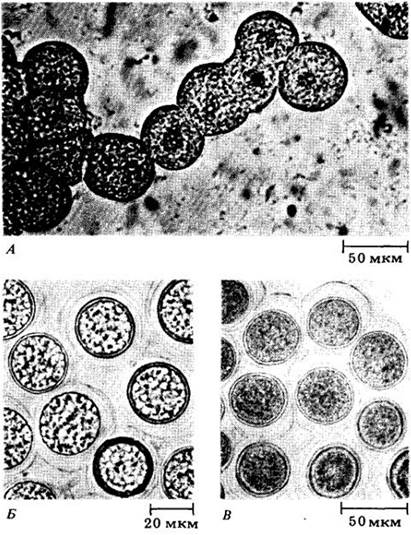
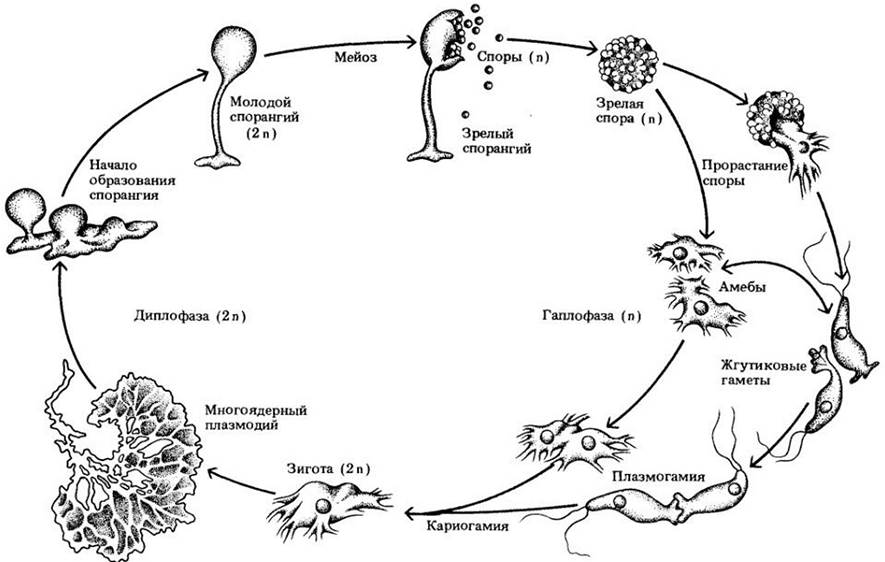
Рост плазмодия продолжается, пока достаточно влаги и пищи. Обычно, если одного из этих факторов не хватает, плазмодий мигрирует из «кормового участка». Тогда его можно заметить пересекающим дороги, лужайки, взбирающимся на деревья и в других самых неожиданных местах. У многих видов прекративший движение плазмодий разделяется на большое число мелких бугорков, одинаковых по объему и размеру, поэтому их образование, вероятно, контролируется химически плазмодием. Каждый бугорок образует зрелый спорангий, обычно на верхушке стебелька; этот спорангий часто имеет сложную форму (рис. 14-11, В, Г). Мейоз происходит в молодых диплоидных спорах после их отделения друг от друга с образованием стенки. При этом возникают четыре ядра, три из которых распадаются, и остается спора с одним гаплоидным ядром. У некоторых представителей миксомицетов отдельный спорангий не образуется, а плазмодий может развиваться либо в плазмодиокарп (рис. 14-11, А), сохраняющий форму плазмодия, либо в эталий (рис. 14-11, Б), т. е. крупный бугорок, являющийся по существу единственным спорангием.
Споры миксомицетов устойчивы к экстремальным условиям внешней среды, некоторые из них способны прорастать после 60-летнего хранения в лаборатории. Таким образом, образование спор обеспечивает здесь не только генетическую рекомбинацию, но и переживание неблагоприятных периодов.
Рис. 14-11. Спорообразующие структуры миксомицетов. А. Плазмодиокарп у Hemitrichia serpula. Б. Эталии Lycogala на коре дерева. В. Спорангий Arcyria cineria. Г. Спорангии Stemonitis splendens
При подходящих условиях споры раскрываются и из них выходит протопласт, остающийся амебоидным или формирующий один-два гладких жгутика. Две эти формы могут легко превращаться друг в друга. Амебы питаются бактериями и органикой, размножаясь митотически. Если источники пищи исчерпаны или условия неблагоприятны, они могут прекращать движение и округляться, выделяя тонкую стенку, т. е. образуя микроцисту. Эти микроцисты сохраняют жизнеспособность в течение года и более и возобновляют активность при улучшении условий.
После периода роста из амеб образуется плазмодий. Этот процесс регулируется целым рядом факторов, включая возраст клеток, условия среды, плотность популяции амеб и химические стимуляторы, роль которых аналогична описанной в случае клеточного слизевика Dictyostelium discoideum (см. с. 124 — 0125). Один из способов образования плазмодия — слияние гамет, которые обычно генетически различны и происходят от разных гаплоидных спор. По существу, это просто амебы или жгутиковые клетки, выступающие в новой роли. У некоторых видов и штаммов плазмодий может возникать и из отдельной амебы; в этом случае он обычно гаплоиден, как и амеба, давшая ему начало.
Жизненный цикл типичного плазмодиального слизевика представлен на рис. 14-12.
Рис. 14-12. Жизненный цикл типичного миксомицета. Половое размножение плазмодиальных слизевиков включает три различные стадии: плазмогамию, кариогамию и мейоз. Плазмогамия заключается в объединении двух протопластов, в результате чего два гаплоидных ядра оказываются в одной клетке. Кариогамия — слияние этих двух ядер, приводящее к образованию диплоидной зиготы и началу так называемой диплофазы жизненного цикла. Плазмодий — это многоядерная, свободно перетекающая масса протоплазмы, способная пройти через шелковую ткань или фильтровальную бумагу практически без изменений. Мейоз возобновляет гаплоидное состояние, и начинается гаплофаза цикла
Отдел Chrysophyta
Хризофиты — это автотрофные, одноклеточные организмы, в изобилии представленные в пресной и морской воде по всему миру. Они содержат хлорофиллы а и с, цвет которых часто маскируется (у золотистых и диатомовых водорослей) вспомогательным пигментом фукоксантином, относящимся к каротиноидам. Запасной углевод хризофитов называется хризоламинарином; часто он накапливается в виде крупного зерна вблизи заднего конца клетки. Клетки хризофитов могут не иметь или иметь клеточную стенку; в последнем случае она образована в основном целлюлозой и покрыта выступающими чешуйками или у многих представителей отдела панцирем из кремнезема. Представители одного из трех классов хризофитов — Xanthophyceae — содержат пигмент вошериксантин, родственный фукоксантину. Организмы этого класса обычно яркого желто-зеленого цвета.
Хлоропласты хризофитов биохимически и структурно аналогичны таковым у бурых водорослей (см. гл. 15) и динофлагеллят. Можно предполагать, что эти органеллы у всех трех групп произошли от одной и той же формы бактерий. До сих пор бактерий с точно такими же признаками не обнаружено, но описанный в 1983 г. в Университете шт. Индиана вид Heliobacterium chlorum, возможно, близок к предшественникам хлоропластов. Это облигатный анаэроб буроватого цвета, способный к азотфиксации. Другое биохимическое сходство хризофитов и бурых водорослей состоит в запасании полисахарида ламинарина (или его более полимеризированной формы хризоламинарина) вне хлоропластов; кроме того, бурые водоросли накапливают маннитол.
Неравные жгутики хризофитов и бурых водорослей сходны (см. рис. 2-26). Высказываются мнения о возможности объединения этих организмов в один отдел.
Хлоропласты динофлагеллят (отдел Pyrrhophyta) биохимически и структурно аналогичны имеющимся у хризофитов и бурых водорослей. Однако динофлагелляты отличаются от этих групп во многих других отношениях, поэтому наличие у них общего предка кажется очень маловероятным. Сходство между тремя отделами, возможно, объясняется тем, что хлоропласты обеих эволюционных линий возникли из одной и той же группы бактерий. Никаких других прямых взаимосвязей между ними нет.
Класс Chrysophyceae
Этот крупнейший класс хризофитов — золотистые водоросли — включает около 500 видов (рис. 14-13). До недавнего времени его считали в основном пресноводной группой, но оказалось, что золотистые водоросли вносят значительный вклад в продуктивность морского планктона, особенно нанопланктона.
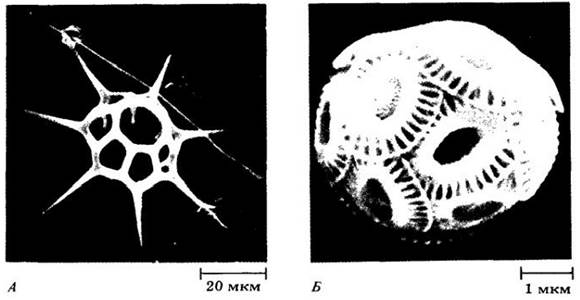
Рис. 14-13. Микрофотографии (сканирующий электронный микроскоп) двух родов морских золотистых водорослей. A. Distephanus speculum, вид холодных вод с кремнеземовым скелетом внутри амебоидного протопласта, содержащего множество мелких хлоропластов. Б. Вид рода Gephyrocapsa, включающего исключительно мелких представителей группы, называемой кокколитофориды. Хотя эти организмы довольно обильны в нанопланктоне, они настолько малы, что их невозможно поймать стандартной планктонной сетью. Они также растворяются в кислых фиксаторах, поэтому их трудно обнаружить и исследовать
Многие золотистые водоросли лишены отчетливой клеточной стенки, но имеют кремнеземовые чешуйки или скелетные элементы, которые могут быть наружными или внутренними и часто сложно структурированы. Большинство представителей этого класса — одноклеточные формы со жгутиками, хотя некоторые жгутиков лишены и иногда сходны с амебами. Эти амебоидные клетки отличаются от корненожек только присутствием хлоропластов, и обе группы, возможно, близкородственны. Некоторые золотистые водоросли действительно захватывают бактерии и другие органические частицы. У видов этой группы в клетке обычно один или два крупных хлоропласта; вблизи заднего конца клетки часто накапливается объемистая хризоламинариновая гранула. Размножение у большинства золотистых водорослей бесполое с помощью зооспор. Некоторые виды образуют колонии.
Класс Xanthophyceae
Организмы этого класса (желто-зеленые водоросли), включающего 550 видов, имеют характерную окраску, давшую название всей группе. Они сходны с золотистыми водорослями присутствием у многих видов хлорофилла с, а отличаются отсутствием фукоксантина. Желто-зеленые водоросли в основном неподвижные существа, хотя некоторые из них имеют амебоидную форму или жгутики, как и их гаметы.
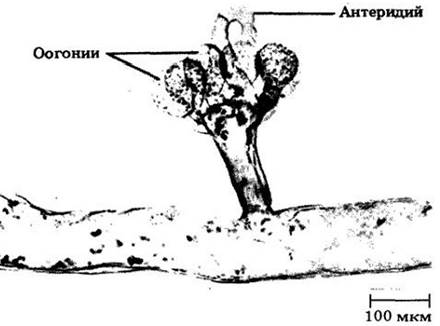
Широко известен в этом классе род Vaucheria («водяной войлок») — ценоцитная, слабо ветвящаяся, нитчатая водоросль. Она размножается как бесполым путем, образуя крупные сложные многожгутиковые зооспоры, так и половым — оогамно (рис. 14-14). Vaucheria распространена в пресных, солоноватых и морских водах. Часто ее можно обнаружить на периодически затопляемом прибрежном иле.
Рис. 14-14. Vaucheria («водяной войлок») — ценоцитный нитчатый представитель хризофитов. Vaucheria — оогамный организм, образующий оогонии и антеридии. Антеридий, показанный на рисунке, пуст
Класс Bacillariophyceae
Эти организмы, называемые диатомовыми водорослями (диатомеями), в большинстве своем одноклеточные. Они являются исключительно важным компонентом фитопланктона и главным источником пищи для пресноводных и морских животных (рис. 14-15). Предполагают, что этот класс объединяет около 5600 живущих ныне видов. Если учитывать и вымершие формы, то диатомовые насчитывают по крайней мере 40 000 видов, причем их видовое разнообразие иногда огромно на очень малом пространстве. Например, в двух мелких образцах ила из океана вблизи Бьюфорта (Северная Каролина) было определено 369 видов. Большинство диатомовых входит в состав планктона, но некоторые обитают на дне, других водорослях или растениях как в пресной, так и в морской воде.
Рис. 14-15. Участок пушистого скопления, образованного переплетенными цепочками диатомовых двух видов рода Rhizosolenia. Скопления такого рода часто обильны в бедных питательными веществами океанических водах; водоросли, представленные на снимке, получены из центра северной части Тихого океана. В вакуолях диатомей в большом количестве обитают симбиотические азотфиксирующие бактерии. Такое сосуществование вносит очень важный вклад в продуктивность вод, бедных питательными веществами

Диатомовые отличаются от других хризофитов отсутствием жгутиков (исключение — мужские гаметы некоторых видов) и уникальными панцирями, или клеточными стенками. Эти тонкие двустворчатые панцири образованы полимерным опаловым кремнеземом (SiO2 х nН2O), причем створки вставляются одна в другую. Тонкая орнаментация панцирей, используемая для определения видов диатомей, традиционно применяется микроскопистами для проверки качества оптических приборов. Электронная микроскопия показала, что тонкие линии на этих панцирях на самом деле образованы большим числом мельчайших, замысловатой формы впадин, пор или канальцев, связывающих живую протоплазму внутри раковин с внешней средой (рис. 14-16). Наиболее характерная особенность протопласта диатомей — буроватые пластиды, содержащие хлорофиллы а и с, а также фукоксантин. Размножение диатомовых в основном бесполое — путем деления клетки (рис. 14-17).
Рис. 14-16. А. Некоторые морские диатомовые под световым микроскопом. Сделанные в сканирующем электронном микроскопе фотографии створки панциря Entogonia (Б) и центрической диатомеи Thalassiosira nordenskioeldii (В). Г. Пеннатная диатомея Pinnularia под световым микроскопом
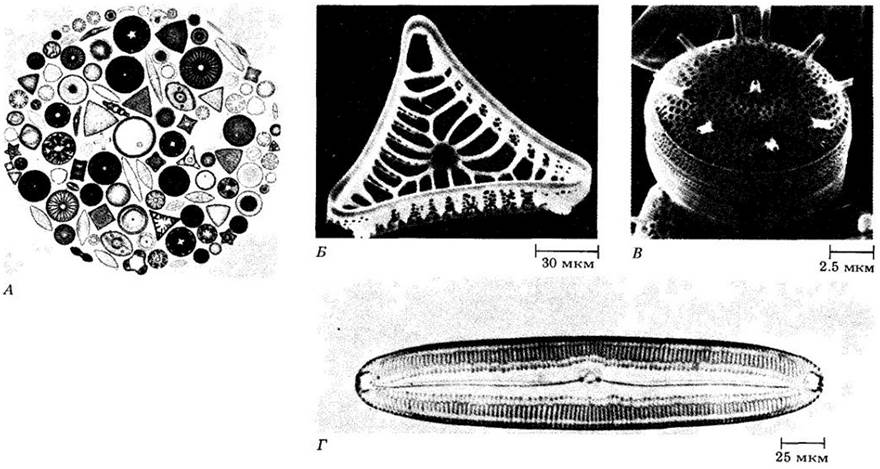
Рис. 14-17. Размножение диатомовых в основном бесполое — путем деления клетки. Каждая дочерняя клетка (слева внизу) получает одну из створок родительского панциря (справа внизу) и достраивает вторую. Родительская створка всегда образует большую половину («крышку») кремнеземовой «коробки», охватывая с боков новую. Таким образом, одна клетка из каждой дочерней пары, как правило, меньше родительской. У некоторых видов панцири увеличиваются за счет разрастания протоплазмы внутри них, но у других они более жесткие. Когда во втором случае особи уменьшатся в размере до 30% максимального диаметра, возможно их половое размножение. Одни клетки функционируют как мужские гаметангии. образуя в процессе мейоза четыре спермия. Другие играют роль женских гаметангиев; три продукта мейоза в них погибают, и образуется только одна яйцеклетка. После сингамии (оплодотворения) возникает ауксоспора, или зигота, которая растет до максимального, характерного для вида размера. Стенки ауксоспор часто резко отличаются от стенок особей того же вида, размножающихся бесполым путем. Созрев, ауксоспора делится, образуя новые панцири, идентичные исходным по всем деталям рисунка. На рисунке показано размножение центрической диатомеи
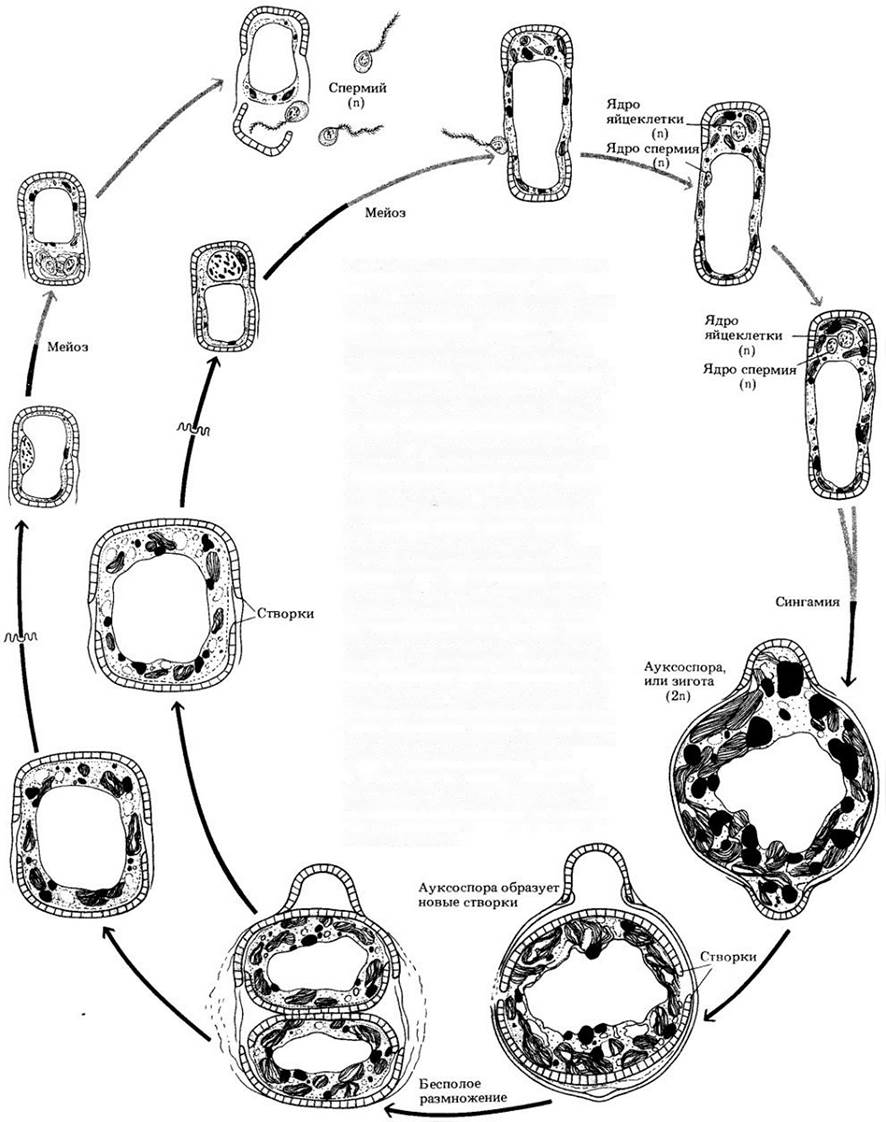
На основе симметрии различают два типа диатомовых: пеннатные (двусторонне симметричные, рис. 14-16, Г) и центрические (радиально симметричные, рис. 14-16, B) формы. Центрические диатомеи наиболее многочисленны в морской воде. Половой процесс у них (если присутствует) представлен оогамией. Мужские гаметы могут иметь один перистый жгутик (единственные жгутиковые клетки в жизненном цикле диатомовых). У пеннатных диатомей половое размножение относится к изогамии, причем обе гаметы не несут жгутиков.
Хотя большинство видов диатомовых автотрофы, некоторые из них могут становиться гетеротрофами и существовать, поглощая органический углерод. Это в основном пеннатные диатомеи, обитающие на относительно мелких участках морского дна. Несколько видов — облигатные гетеротрофы, лишенные хлорофилла и не способные к фотосинтезу. С другой стороны, некоторые диатомовые, без характерных для класса панцирей, живут симбиотически в крупных морских простейших (фораминиферах), обеспечивая своего хозяина органическим углеродом.
Несмотря на отсутствие жгутиков и других локомоторных органелл, многие виды пеннатных диатомей подвижны. Это движение связано с тонко регулируемой секрецией веществ в ответ на самые разнообразные физические и химические воздействия. По-видимому, вдоль средней оси створок всех подвижных диатомовых проходит так называемый шов, который по существу является двумя порами, связанными с тонкой, сложно устроенной щелью в панцире. Многие неподвижные диатомеи прикрепляются друг к другу своими панцирями, образуя длинные нити (рис. 14-15). Шов как локомоторное приспособление, вероятно, возник в процессе эволюции путем изменения апикальных пор, выделяющих вещества для скрепления неподвижных диатомовых в такие нити.
Диатомея движется в ответ на внешние механические раздражения, свет, тепло, токсины с помощью примыкающих к шву сократительных пучков. Их сокращение вызывает движение обезвоженных кристаллических тел в область резервуаров вблизи пор шва. Отсюда эти кристаллические тела выбрасываются в поры. Здесь они поглощают воду и набухают, превращаясь в скрученные фибриллы. Фибриллы движутся вдоль шва, пока не соприкоснутся с какой-либо поверхностью, немедленно прилипают к любому субстрату и затем сокращаются. Если объект, к которому они прикрепились, достаточно крупный, то диатомея притягивается к нему, оставляя след секретируемого вещества, подобно улитке, которая выделяет слизь. Если объект мал, он перемещается вдоль шва, а диатомея остается на месте. Однако подвижные диатомовые обычно находятся в покое. Они могут передвигаться лишь на ограниченное расстояние, так как в каждый момент времени запас кристаллических тел, необходимых для локомоции, лимитирован.
Кремнеземовые панцири диатомей накапливались миллионы лет, образуя мелкозернистый рыхлый порошок, называемый диатомовым илом, который используется как абразивный материал для тонкой полировки, а также для фильтрования и выделения веществ. Считается, что в 1 см3 диатомового ила около 4,6 млн. таких панцирей. В нефтяном бассейне Санта-Мария (Калифорния) подземные отложения диатомового ила достигают мощности 900 м, а вблизи Ломпока (Калифорния) его ежегодно добывается для промышленных целей 270 000 т.
Диатомовые становятся обильными в отложениях возрастом около 100 млн. лет, т. е. в меловом периоде. Многие из ископаемых видов идентичны современным, что указывает на необычную стабильность представителей этого класса на протяжении геологической истории.
Отдел Pyrrhophyta
Большинство этих организмов, называемых динофлагеллятами, — одноклеточные двужгутиковые формы (см. табл. 14-1). К ним относится более 1000 видов, многие из которых многочисленные и высокопродуктивные компоненты морского планктона; имеются и пресноводные представители. У динофлагеллят жгутики проходят в желобках, один из которых пояском окружает тело, а другой расположен перпендикулярно первому. Биение жгутиков в желобках заставляет особь кружиться во время движения «волчком». Опоясывающий жгутик имеет лентовидную форму. Встречаются также многочисленные неподвижные формы, иногда без жгутиков. Некоторые роды динофлагеллят захватывают твердые пищевые частицы, частично или полностью получая таким способом необходимое питание.
Многие динофлагелляты имеют причудливую форму с плотными целлюлозными пластинками, образующими клеточную стенку (теку), часто напоминающую странный шлем или часть древних доспехов (рис. 14-18, 14-19). Пластинки этой стенки находятся в вакуолях под плазматической мембраной, а не снаружи ее, как клеточная стенка большинства водорослей.
Рис. 1418. «Доспехи» некоторых динофлагеллят состоят из целлюлозных пластинок, лежащих в вакуолях под плазматической мембраной. Эти вакуоли у родов, кажущихся «голыми», могут содержать или не содержать целлюлозные пластинки
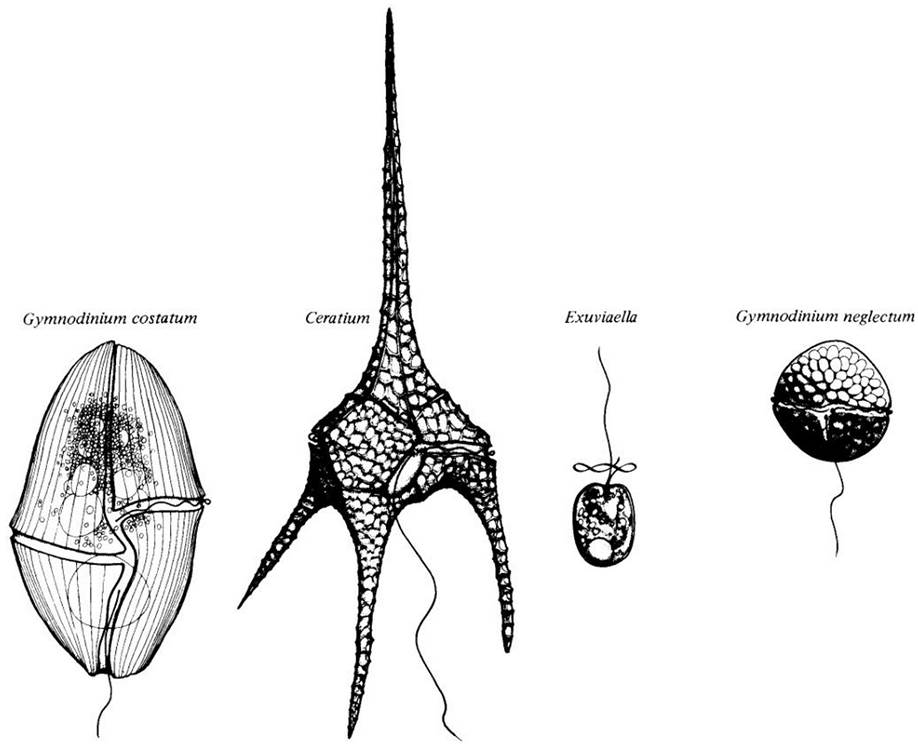
Рис. 14-19. A. Ceratium triops, динофлагеллята с причудливым панцирем. Б. Noctiluca scintillans, биолюминесцентная морская динофлагеллята. R. Gonyaulax polyedra, динофлагеллята, ответственная за появление живописных «красных приливов» у берегов южной Калифорнии
Динофлагелляты, как правило, содержат хлорофиллы а и с, которые обычно маскируются каротиноидами, в том числе близким к фукоксантину перидином. Их хлоропласты, как уже упоминалось выше, вероятно, произошли от той же группы бактерий, что и у хризофитов и бурых водорослей, но во всех других отношениях они резко отличаются от представителей этих отделов.
Запасной углевод динофлагеллят — крахмал. Некоторые виды нс содержат хлорофилла и являются гетеротрофами, но их структура четко указывает на связь с другими представителями этого отдела. Даже автотрофы обычно сильно нуждаются в витамине В12 (как и многие диатомовые), т. е. их автотрофность требует по крайней мере этой оговорки. Некоторые динофлагелляты способны заглатывать другие клетки. Предполагают, что бесцветные формы добывают пищу именно таким способом, а также поглощая мелкие частицы органики.
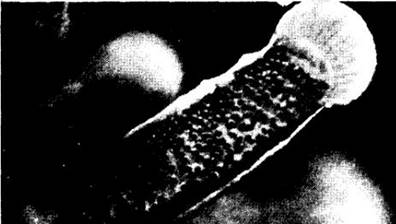
Цианобактерии — обычные симбионты динофлагеллят, которые в свою очередь вступают в симбиоз со многими другими организмами, включая губок, медуз, актиний, оболочников, кораллы, осьминогов и кальмаров, брюхоногих, турбеллярий и некоторых протистов. У гигантских двухстворчатых моллюсков из семейства Tridachnidae дорсальная часть внутренней поверхности мантии может быть темно-шоколадной из-за присутствия симбиотических динофлагеллят. Выступая в роли симбионтов, они теряют теку и выглядят как золотистые сферические клетки, называемые зооксантеллами (рис. 14-20).
Рис. 14-20. Зооксантеллы — симбиотическая форма динофлагеллят, представленная здесь в щупальце коралла — вносят существенный вклад в продуктивность коралловых рифов
Зооксантеллы в основном обеспечивают фотосинтетическую продуктивность, дающую возможность расти коралловым рифам в тропических водах, весьма бедных питательными веществами. Ткани кораллов могут содержать до 30 000 симбиотических динофлагеллят в 1 мм3, главным образом внутри клеток, выстилающих пищеварительную полость полипа. Поскольку водорослям для фотосинтеза необходим свет, содержащие их кораллы, как правило, растут в океане на глубинах не более 60 м. Многие особенности строения кораллов связаны со светоулавливающими свойствами различных геометрических конструкций. Аналогичное явление можно наблюдать в кронах деревьев, характер ветвления которых должен обеспечить максимальную освещенность листьев солнцем.
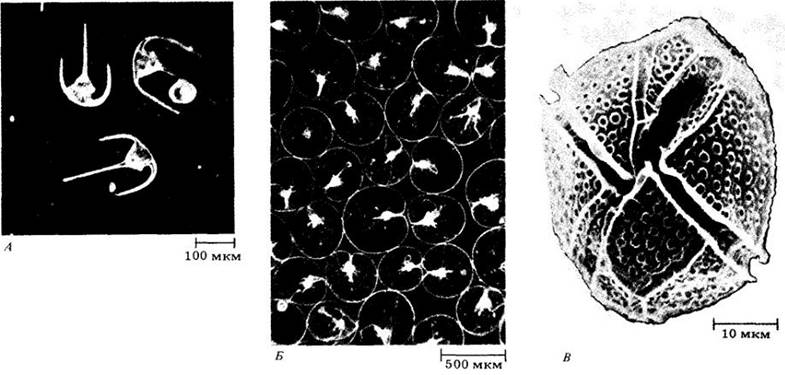
Динофлагелляты играют также важную роль в жизни человека. Зимой и весной 1974 г. западное побережье Флориды было опустошено мощным «красным приливом», 25-м по счету с 1844 г. Сотни тысяч мертвых рыб покрыли пляжи; были потеряны миллионы долларов, получаемых от туризма. Такие явления вызываются необычными вспышками численности динофлагеллят, придающими морской воде красный или бурый цвет («цветение» воды) (рис. 14-21). Эти динофлагелляты поедаются не только рыбой, которая может получать непосредственные отравления, но и двустворчатыми моллюсками, в частности мидиями, обычно от этого не страдающими. Моллюски накапливают и концентрируют токсины, синтезируемые водорослями, и в зависимости от характера яда представляют опасность для людей, потребляющих их в пищу. Осенью 1972 г. «красный прилив» впервые наблюдался у побережья Новой Англии от шт. Мэн до залива Кейп-Код. После этого 26 человек отравились моллюсками, питавшимися Gonyaulux excavata. Общественность была настолько потрясена, что промысел моллюсков в шт. Массачусетс упал в следующие четыре года на треть. «Красные приливы» возникают периодически. Во время самого мощною из них после 1972 г. на всем побережье шт. Мэн добыча моллюсков была официально запрещена с середины августа до середины октября 1980 г. Пищевая промышленность потеряла на устрицах, мидиях и других двустворках 7 млн. долларов.
Рис. 14-21. Ptychodiscus brevis, динофлагеллята без панциря, вызывающая «красные приливы» у побережья Флориды. Изогнутый поперечный жгутик лежит в бороздке, опоясывающей организм. Продольный жгутик (видна только его часть) проходит от центра клетки вниз налево. Апикальное углубление — отличительный признак рода Ptychodiscus

Яды, образуемые некоторыми динофлагеллятами, например, Gonyaulax catenella, являются необычайно мощными нервными токсинами. Химическая природа и биологическая активность большинства из них относительно хорошо изучены. С другой стороны, причины появления «красных приливов» понятны плохо. Концентрации питательных веществ и некоторых следовых металлов, сточные воды, соленость и температура воды, ветры, свет и многие другие факторы, вероятно, играют в этом некоторую роль. Периодичность их возникновения дополняется следующим наблюдением: если во время «цветения» воды условия меняются к худшему, динофлагелляты могут терять жгутики, образуя покоящиеся цисты, которые оседают на дно и пребывают в состоянии покоя до тех пор, пока условия вновь не станут благоприятными.
Основной способ размножения динофлагеллят — продольное деление, когда каждая дочерняя клетка получает один из жгутиков и часть теки, а затем в весьма сложной последовательности достраивает недостающие компоненты (см. «Митоз у динофлагеллят», с. 231). Некоторые неподвижные формы образуют зооспоры. Иногда типично динофлагеллятную структуру у них имеют только зооспоры, в то время как зрелые особи лишены жгутиков и могут объединяться друг с другом в нити. Для ряда динофлагеллят описан и половой процесс. Это в основном изогамия; иногда анизогамия.
Отдел Euglenophyta
Известно более 800 видов эвгленовых, живущих в основном в пресной воде, особенно в воде, обогащенной органикой (табл. 14-1). Длина их колеблется от 10 до 500 мкм, а форма самая разнообразная. Все они одноклеточные, кроме колониального рода Colacium.
Как уже говорилось, сходство хлоропластов эвгленовых и зеленых водорослей позволяет предполагать, что органеллы фотосинтеза произошли независимо в обеих группах от одной формы бактерий, возможно, сходной с Prochloron (см. рис. 11-2, 11-17). В других отношениях эти два отдела резко различаются. Среди примерно 40 родов эвгленовых около трети имеет хлоропласты, содержащие хлорофиллы а и b и несколько каротиноидов.
Эвгленовые запасают углеводы в форме полисахарида парамилона, не обнаруженного больше ни в одной группе организмов. Это запасное вещество, как у всех автотрофов, кроме зеленых водорослей и растений, образуется вне хлоропластов. Среди эвгленовых, лишенных хлоропластов, некоторые абсорбируют органическое вещество, другие его заглатывают. Как уже упоминалось, представители этого отдела по существу являются жгутиковыми простейшими, которые приобрели в ходе эволюции хлоропласты; возможно, обе группы следует объединить.
Эвгленовые размножаются делением, в ходе которого отдельные клетки сохраняют подвижность. Ядерная оболочка при митозе не разрушается, как и у большинства зеленых водорослей, динофлагеллят, многих грибов и некоторых инфузорий. Центриоли функционируют как базальные тельца и образуют обычное веретено деления под ядерной оболочкой. Хромосомы остаются конденсированными в течение интерфазы и митоза, как у динофлагеллят. Половое размножение неизвестно.
Название отдела производится от широко распространенного рода Euglena (эвглена), многие виды которого имеют удлиненную форму (рис. 14-22). Их клетка сложно организована и содержит большое число мелких хлоропластов. На ее конце отходят длинный, хорошо заметный жгутик с очень тонкими волосками и не выходящий из особого углубления (см. ниже) короткий жгутик. Длинный жгутик обычно находится впереди клетки, как леска спиннинга.
Рис. 14-22. A. Euglena; видны два запасных парамилоновых тельца и природа пелликулы. Б. Структура Euglena по данным электронной микроскопии
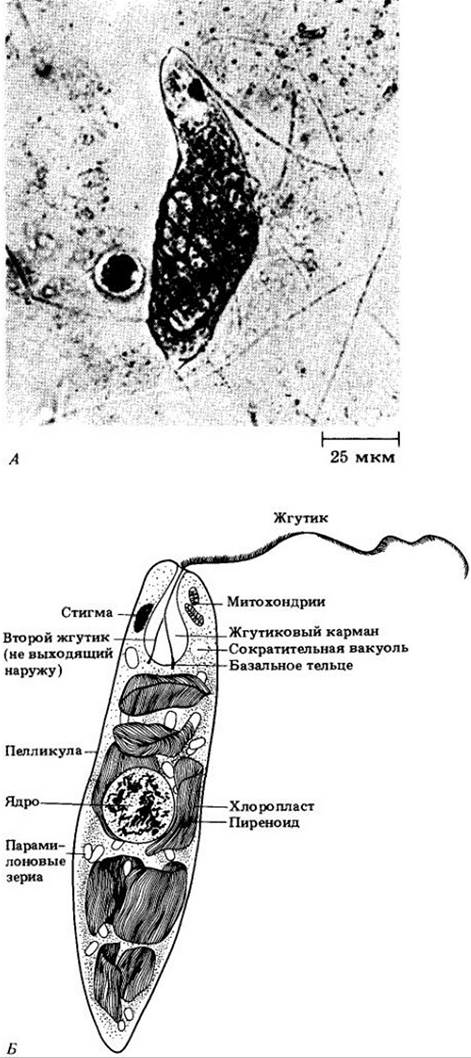
У эвглены жгутики прикреплены к основанию колбовидного жгутикового кармана (или глотки) на переднем конце клетки. Сократительная вакуоль собирает избыток воды из всех ее частей и выбрасывает ее в этот же карман. Клетка ограничена плазматической мембраной, под которой проходит по спирали несколько гибких, соединенных друг с другом белковых тяжей. Вместе с мембраной они образуют структуру, называемую пелликулой. Гибкая пелликула в отличие от жесткой клеточной стенки растений позволяет эвглене изменять форму, обеспечивая альтернативный способ локомоции для обитающих в иле форм.
Если культуру эвглены поместить вблизи окна, через которое падают солнечные лучи, то в воде образуется хорошо заметное зеленое скопление клеток, движущееся по мере изменения освещенности всегда в сторону самого светлого (но не слишком светлого) участка. Если свет очень яркий, эвглены его избегают. Их способность ориентироваться по свету, возможно, обусловлена наличием двух особых структур — стигмы и связанного с ней фоторецептора, представленного вздутием в основании жгутика. В зависимости от положения особи по отношению к свету различное «затенение» фоторецептора пигментированной стигмой, вероятно, определяет направление движения. Среди бесцветных эвгленовых стигму имеют немногие.
Некоторые виды эвглен могут жить в темноте, не фотосинтезируя, если они обеспечены источником углерода, необходимыми витаминами и минеральными веществами. Если некоторые штаммы держать на свету при соответствующей температуре и в богатой пищей среде, клетки могут удваиваться быстрее, чем хлоропласты, с образованием не способных к фотосинтезу особей, тем не менее неограниченно долго существующих в подходящей среде. По сути дела, эвгленовые — это простейшие с нестабильными хлоропластами.