СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 17. СПОРОВЫЕ СОСУДИСТЫЕ РАСТЕНИЯ
Репродуктивные системы
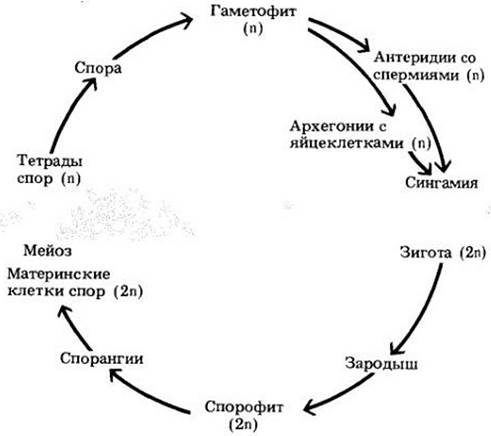
Для всех сосудистых растений типичны оогамия и чередование поколений, при котором гаметофит зависит в отношении питания от спорофита (рис. 17-6). Очевидно, что оогамия наиболее подходит для наземных растений, так как при ней перемещаться в чуждой среде за пределами растения должен только один тип гамет, а не оба. В жизненном цикле сосудистых растений доминирует спорофит; он крупнее и намного структурно сложнее гаметофита. У моховидных, как говорилось в гл. 16, гаметофит почти всегда крупнее спорофита, хотя, как и у сосудистых растений, он обычно проще устроен.
Рис. 17-6. Обобщенный жизненный цикл сосудистого растения
Гомоспория и гетероспория
Ранние сосудистые растения давали только один тип спор; их называют гомоспоровыми (равноспоровыми). Среди современных сосудистых гомоспория встречается у псилотов, хвощей, некоторых плауновидных и почти у всех папортников. В результате мейоза у гомоспоровых возникает только один тип спор, которые прорастают в обоеполые, т. е. имеющие и антеридии, и архегонии, гаметофиты.
Гетероспория (разноспоровость) — образование двух типов спор в двух разных типах спорангиев — встречается у некоторых плауновидных, немногих папоротников и у всех семенных растений. В ходе эволюции неродственных групп сосудистых растений она возникала неоднократно и была обычной уже в девонском периоде, т. е. более 360 млн. лет назад. Споры при этом называются микро- и мегаспорами и образуются соответственно в микро- и мегаспорангиях. Хотя «микро» указывает на малые, а «мега» на крупные размеры, мегаспоры нс всегда крупнее микроспор, особенно у семенных растений. Однако между ними всегда существуют функциональные различия. Микроспоры дают начало мужским гаметофитам (микрогаметофитам), а мегаспоры — женским (мегагаметофитам). Оба этих типа однополых гаметофитов гораздо мельче гаметофитов гомоспоровых сосудистых растений. У гетероспоровых они развиваются под оболочкой споры, а у гомоспоровых — вне ее.
У хвощей все споры, будучи морфологически идентичными, неодинаковы в физиологическом отношении — некоторые из них дают начало однополым гаметофитам. Можно считать поэтому, что у хвощей наблюдается физиологическая гетероспория, являющаяся, вероятно, ступенью в эволюции только что рассмотренной морфологической гетероспории.
Гаметофиты и гаметы
Относительно крупные гаметофиты равноспоровых растений независимы от спорофита в своем питании, хотя подземные гаметофиты некоторых видов, например, псилотов и ряда плаунов, гетеротрофны и связаны питанием с эндомикоризными грибами. Но у других плаунов, как и у большинства папоротников и хвощей, гаметофиты свободноживущие и фотосинтезирующие. В противоположность этому гаметофиты разноспоровых получают питательные вещества от спорофита.
Эволюция гаметофита сосудистых растений характеризуется постепенным уменьшением размера и сложности строения, причем наиболее заметна эта редукция у покрытосеменных (см. с. 332). Архсгонии имеются у всех споровых сосудистых растений и у большинства голосеменных, но отсутствуют у всех покрытосеменных; антеридиев, характерных для споровых, у всех голосеменных и покрытосеменных нет. У споровых, включая папоротники, подвижный спермий плывет в водной среде к архегонию, следовательно, эти растения должны расти в таких местообитаниях, где вода, по крайней мере периодически, в изобилии.
Семя — уникальная структура, в которой зародыш сбрасывается с родительского растения. При этом он окружен прочной оболочкой вместе с запасом пищи, способствующим его приживанию. Семя появилось в ряде эволюционных линий сосудистых растений по крайней мере 360 млн. лет назад. Среди этих групп — плауновидные и хвощевидные, которые обсуждаются в данной главе, посвященной споровым, поскольку все современые представители этих отделов семян не имеют. Эволюция семени — одного из главных факторов, обусловивших господствующее положение семенных растений, и основные черты таких растений, будут рассмотрены в гл. 18.