СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 17. СПОРОВЫЕ СОСУДИСТЫЕ РАСТЕНИЯ
Отделы споровых сосудистых растений
Три отдела этих растений — риниофиты (Rhyniophyta), зостерофиллофиты (Zosterophyllophyta) и тримерофиты (Тrimerophyta) — процветали в девонский период и вымерли к его концу, т. е. около 360 млн. лет назад, причем риниофиты появились по крайней мере 420 млн. лет назад, в позднем силуре. Все это относительно простые по структуре организмы. Четвертый отдел споровых Progymnospermophyta, или прогимноспермы, будет рассматриваться в гл. 18, так как его представители, возможно, были предками голосеменных и покрытосеменных (рис. 17-7). Кроме вымерших растений мы обсудим в этой главе отделы Рsilotophyta, Lycophyta, Sphenophyta и Pterophyta, в которых имеются ныне живущие представители.
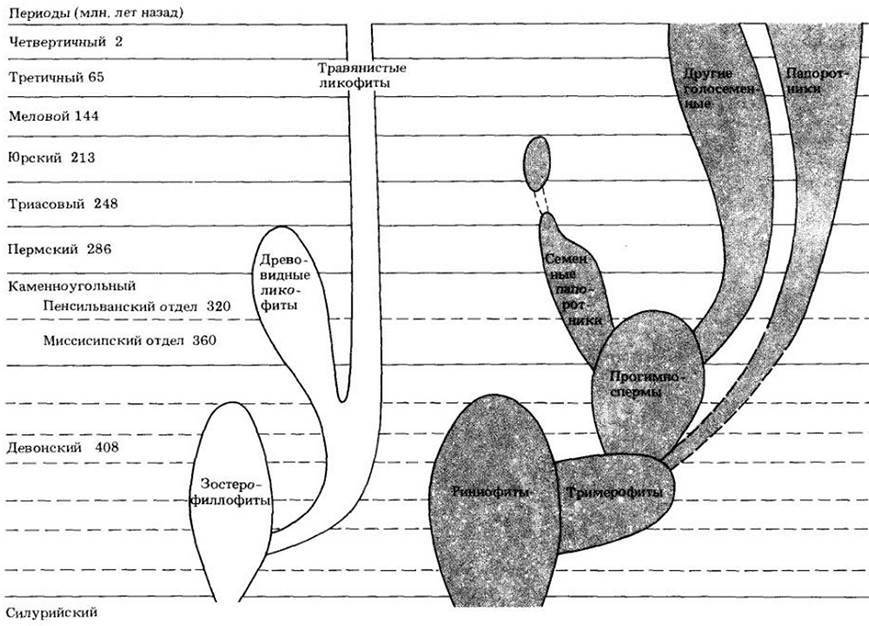
Рис. 17-7. Возможный ход эволюции сосудистых растений, начиная от зостерофиллофитов и риниофитов. Продолжительность девонского периода в общем масштабе временной шкалы увеличена, не показаны связи с цветковыми растениями. Не ясно, от кого произошли псилотофиты и хвощевидные. Описано всего несколько родов ископаемых растений, являющихся, возможно, промежуточными между риниофитами, появившимися в силурийском периоде (около 420 млн. лет назад), и
зостерофиллофитами, древнейшие остатки которых примерно на 10 млн. лет моложе. Точную природу взаимосвязей между этими двумя группами и их общего предка еще предстоит установить
Общую модель нарастания разнообразия флоры можно представить, как поочередное наступление господства четырех крупных групп, сопровождающееся резким вытеснением предыдущих доминантов. В каждом случае группы, захватывая ведущее положение, достигали высокого видовою богатства. Основное участие в этом историческом процессе принимали:
1. Первые сосудистые растения с простой и, вероятно, исходной для прочих групп морфологией. К ним относятся риниофиты, зостерофиллофиты и тримерофиты (рис. 17-8). Эти примитивные формы доминировали с позднего силура до среднего девона, т. е. примерно 420 — 370 млн. лет назад (см. рис. 17-1).
2. Папоротники, плауновидные, хвощевидные, прогимноспермы. Эти более сложно устроенные растения господствовали с позднего девона до конца каменноугольного периода (см. рис. 17-9, 18-1), примерно 380 — 290 млн. лет назад. Приложение к главе дает представление о растениях той эпохи (см. с. 307).
3. Семенные растения, появившиеся в позднем девоне (по крайней мере, 360 млн. лет назад) и к началу пермскою периода уже представленные многими эволюционными линиями. Голосеменные преобладали во флорах суши на протяжении большей части мезозойской эры, примерно до 100 млн. лет назад.
4. Цветковые растения, древнейшие остатки которых имеют возраст около 127 млн. лет. Они завоевали сушу за 20 — 30 млн. лет и с тех пор сохраняют господствующее положение.
Отдел Rhyniophyta
К этому отделу принадлежат самые древние из детально известных нам сосудистых растений. Они появились в позднем силуре (не менее 420 млн. лет назад) и вымерли в среднем девоне (около 380 млн. лет назад). Еще более древние сосудистые растения, ископаемые остатки, которых по крайней мере на 15 млн. лет старше, были, вероятно, сходны с риниофитами. Отдел включает споровые растения, состоящие из простых, дихотомически ветвящихся осей с верхушечными спорангиями. Дифференцировка на корни, стебли или листья отсутствовала, виды были равноспоровыми. Название отдела происходит от деревни Райни1 в Шотландии, близ которой остатки этих примитивных растений прекрасно сохранились в кремнистых породах.
1По-латыни ее название (Rhynie) читается «Риниэ». — Прим. ред.
Куксония (Cooksonia), представитель риниофитов, обитавший, как полагают, на заболоченных низинах, — древнейшее из известных нам сосудистых растений (см. рис. 1-5, 17-1). Ее тонкие безлистные надземные стебли достигали в длину 6,5 см; спорангии были шаровидными. Хотя о нижних частях этих растений ничего не известно, вполне вероятно, что куксония имела корневище, т. е. подземный стебель, от которого отходили надземные ветви. В мацерированных кусочках осей обнаружены трахеиды. Куксония вымерла к раннему девону, около 390 млн. лет назад.
Наиболее известный представитель риниофитов — риния (Rhynia) (рис. 17-8, A), вероятно болотное растение, безлистные, дихотомически ветвящиеся стебли которого отходили от корневища с пучками водопоглощающих ризоидов. Надземные стебли ринии длиной 20 — 50 см и толщиной 3 — 6 мм были покрыты кутикулой, имели устьица и служили фотосинтезирующими органами.
Рис. 17-8. Древние сосудистые растения. А. Риниофит Rhynia major — одно из наиболее простых известных нам сосудистых растений. Его безлистный стебель дихотомически ветвился. Герминальные спорангии высвобождали споры, вероятно, за счет продольного расщепления. Б. У рода Zosierophyllum и других зостерофиллофитов собранные в терминальные колоски спорангии расщеплялись по определенным бороздам вокруг своего дистального кран. Более крупные, чем риниофиты, эти растения, как и последние, как правило, дихотомически ветвились, были голыми, шиповатыми или покрытыми зубчиками. R. Тримерофиты — еще более крупные и сложнее устроенные растения с крепкой центральной осью и боковыми ветвями меньшего размера, которые дихотомически ветвились и часто заканчивались группами парных, суженных к концам спорангиев. Наиболее известные роды из этой группы — Psilophyton и Trimerophyton. На рисунке приведена реконструкция вида Psiíophyton princeps. Отдельные экземпляры Rhynia major достигали в высоту 0,5 м, а некоторые тримерофиты — более 1 м (см. также рис. 17-1)
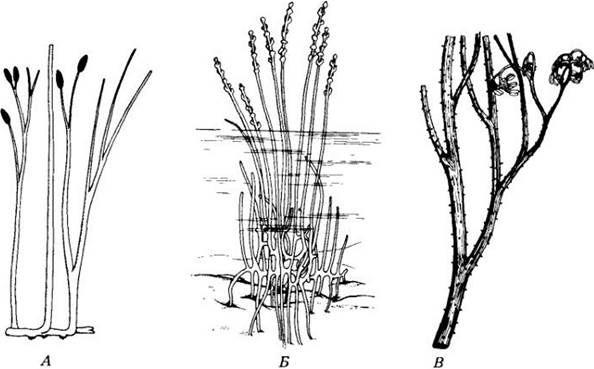
Рис. 17-9. Реконструкция болотистого леса каменноугольного периода (см. также рис. 18-1)

Рис. 17-10. Psilotum nudum. А. Поперечный разрез стебля. показывающий сформировавшиеся ткани. Б. Протостела; видны ксилема и флоэма
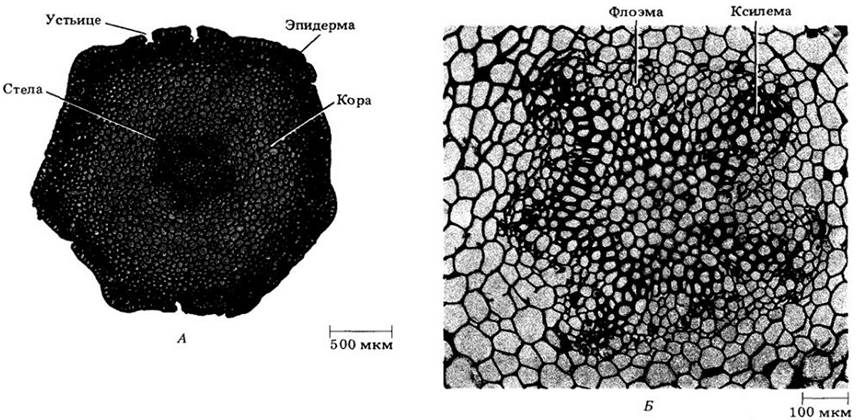
Рис. 17-11. Подземный гаметофит Psilotum nudum. Гаметофиты обоеполые, т. е. несут и антеридии, и архегонии

Рис. 17-12. A. Tmesipteris parva на стволе древовидного папоротника Cyathea australis в Новом Южном Уэльсе, Австралия. Б. Tmesipteris lanceolate из Новой Каледонии
Рис. 17-13. Жизненный цикл псилота. У этого рода из корневищ вырастают дихотомически ветвящиеся надземные побеги. На них в пазухах чешуевидных выростов группами по три развиваются спорангии. Для псилота, как и для всех сосудистых растений, характерно чередование гетероморфных поколений с доминированием спорофита. В результате мейоза внутри спорангия образуются споры, и начинается гаметофитная фаза. Псилот равноспоровый; его споры дают начало обоеполым, т. е. несущим одновременно антеридии и архегонии, гаметофитам. Чтобы достичь яйцеклетки, многожгутиковый спермий нуждается в воде. При сингамии образуется зигота, из которой развивается спорофитное поколение. Зародыш формируется в брюшке архегония, причем молодой спорофит получает питание от гаметофита через свою стопу, внедренную в его ткань. В конечном итоге спорофит отделяется от стопы и начинает независимое существование
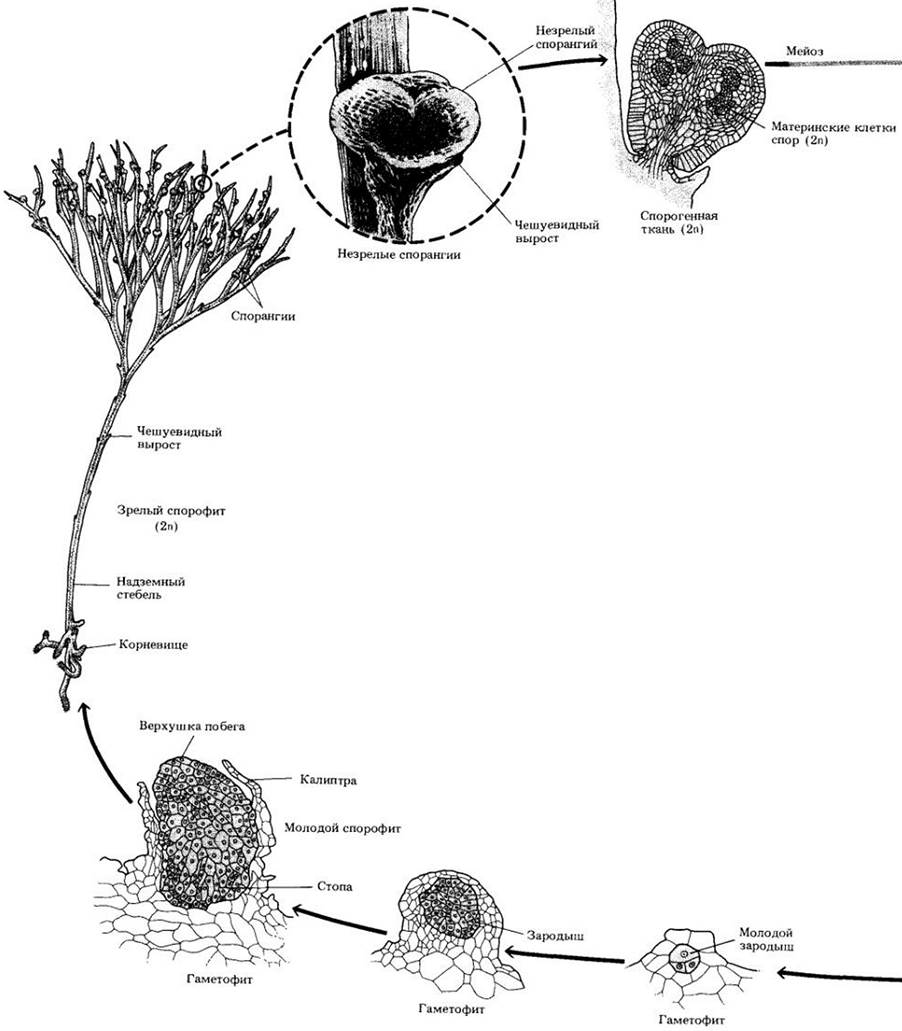
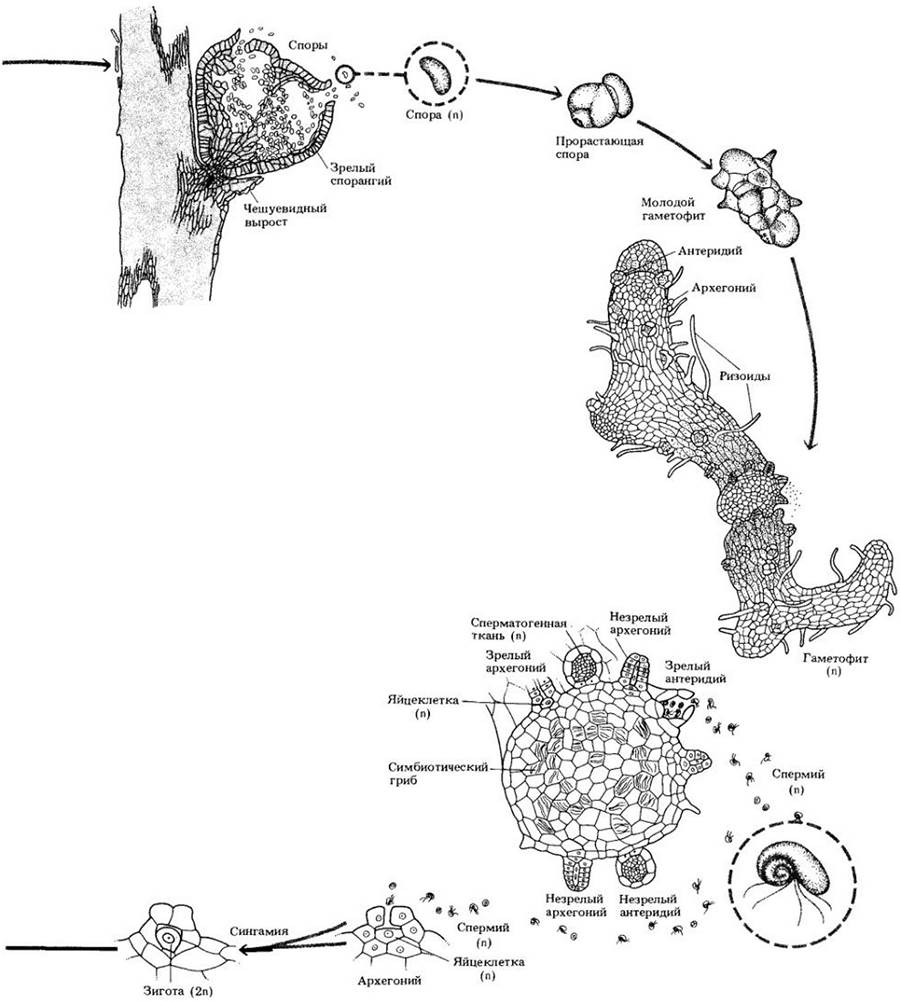
Рис. 17-14. А. Поперечный срез стебля Lycopodium complanatum, на котором видны зрелые ткани. Б. Протостела этого же вида; видны ксилема и флоэма. И стебель, и корень плаунов протостелические (см. также рис. 17-2)
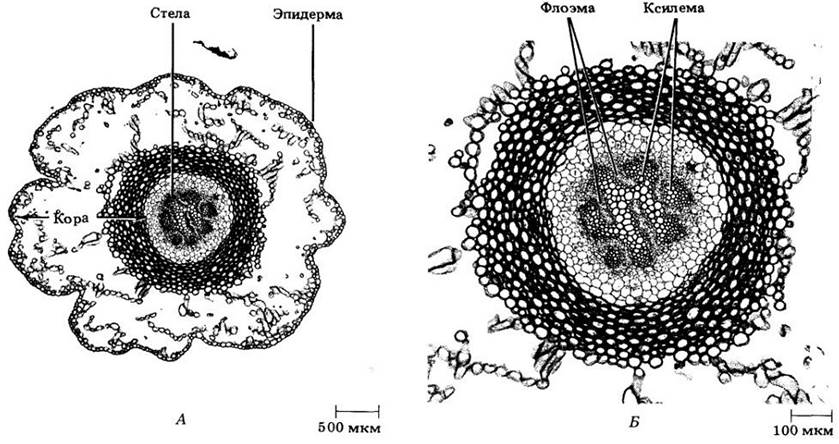
Рис. 17-15. Lycopodium lucidulum —представитель плаунов, не имеющих стробила; спорофиллы рассеяны среди стерильных микрофиллов
По внутренней структуре риния напоминала современные сосудистые растения. Единственный слой поверхностных клеток — эпидерма — окружал ассимиляционную ткань коры, а центр оси состоял из сплошного тяжа ксилемы, окруженного двумя слоями клеток, возможно, флоэмных. По- видимому, первыми развивались клетки ксилемы в центральной части тяжа, а последними — на его периферии.
Рис. 17-16. А. Selaginella kraussiana — стелющееся, ползучее растение. Видны придаточные корни, образующиеся на стеблях. Б. S. rupestris со стробилами. В. S. willdenowii — растение тропиков Старого Света, встречающееся во Вьетнаме, Малайзии и Гималаях. Этот теневыносливый вид с переливчатыми, металлически блестящими голубыми листьями может забираться по деревьям на семиметровую высоту
Отдел Zosterophyllophyta
Ископаемые остатки представителей этого отдела вымерших сосудистых растений находят в ранне позднедевонских отложениях возрастом примерно от 408 до 370 млн. лет. Как и риниофиты, они были безлистными и дихотомически разветвленными. Возможно, растения этой группы обитали в воде. Надземные стебли покрывала кутикула, но только самые верхние ветви имели устьица. Это может свидетельствовать о том, что нижние были погружены в ил. Предполагают, что у зостерофиллума (Zosterophyllum) эти ветви часто давали боковые ответвления, вильчато разделявшиеся на две оси, одна из которых росла вверх, а другая вниз (рис. 17-8, Б). Растущие вниз ответвления, по-видимому, служили опорой растению, позволяя ему распространяться в стороны. Свое название зостерофиллофиты получили из-за их общего сходства с современным покрытосеменным Zostera, растущим в морской воде и внешне напоминающим злаки.
В отличие от риниофитов спорангии зостерофиллофитов, имевшие шаровидную или бобовидную форму, располагались латерально на коротких ножках. Растения были равноспоровыми. Хотя внутренняя структура зостерофиллофитов в основном такая же, как у риниофитов, первыми достигавшие зрелости клетки ксилемы располагались здесь по периферии ксилемного тяжа, а развивавшиеся последними — в его центре.
Можно почти с полной уверенностью считать, что зостерофиллофиты — предки плауновидных. Спорангии древних плауновидных также располагались латерально, и ксилема в обоих отделах развивалась центростремительно (от периферии к центру). Представители двух этих отделов заметно отличаются от риниофитов и тримерофитов.
Отдел Trimerophyta
Этот отдел произошел, вероятно, непосредственно от риниофитов, а его представители, по-видимому, стали предками папоротников, прогимноспермов и, не исключено, также хвощей. Тримерофиты, более крупные и более сложно устроенные по сравнению с риниофитами и зостерофиллофитами растения (рис. 17-8, В), впервые появились в раннем девоне, около 395 млн. лет назад, и вымерли к концу среднего девона, примерно 20 млн. лет спустя, просуществовав, таким образом, относительно короткий промежуток времени.
Хотя тримерофиты более продвинуты в эволюционном отношении, чем риниофиты, они еще лишены листьев. На главной оси формировались системы боковых ветвей, неоднократно разделявшихся дихотомически. Эти растения, подобно риниофитам и зостерофиллофитам, были равноспоровыми. Некоторые из более мелких ветвей заканчивались удлиненными спорангиями, тогда как другие были исключительно вегетативными. Кроме усложнившегося способа ветвления тримерофиты имели и более массивный, чем у риниофитов, проводящий тяж, который вместе с мощной корой, состоявшей из толстенных клеток, вероятно, давал опору довольно крупному растению. Как и у риниофитов, первые клетки ксилемы у тримерофитов формировались в центре (центробежное развитие). Название отдела происходит от греческих слов «tri», «meros» и «phyton» и означает «растение из трех частей». В нем отражено тройчатое разделение вторичных ветвей у рода Trimerophyton.
Рис. 17-17. Жизненный цикл плауна. Это растение, как и псилот, равноспоровое, поэтому их жизненные циклы схожи. В результате мейоза образуются споры, дающие начало обоеполым гаметофитам. Последние у некоторых видов плауна подземные, и для их нормального роста, как показано на рисунке, требуется присутствие микоризного гриба. Чтобы произошло оплодотворение, необходима вода, в которой двужгутиковые спермии плывут к архегонию. При сингамии возникает зигота. Развитие зародыша идет внутри брюшка архегония. Молодой спорофит, иногда длительное время прикрепленный к гаметофиту, в конце концов становится независимым. Спорофиллы у многих видов плаунов собраны, как видно на рисунке, в стробилы
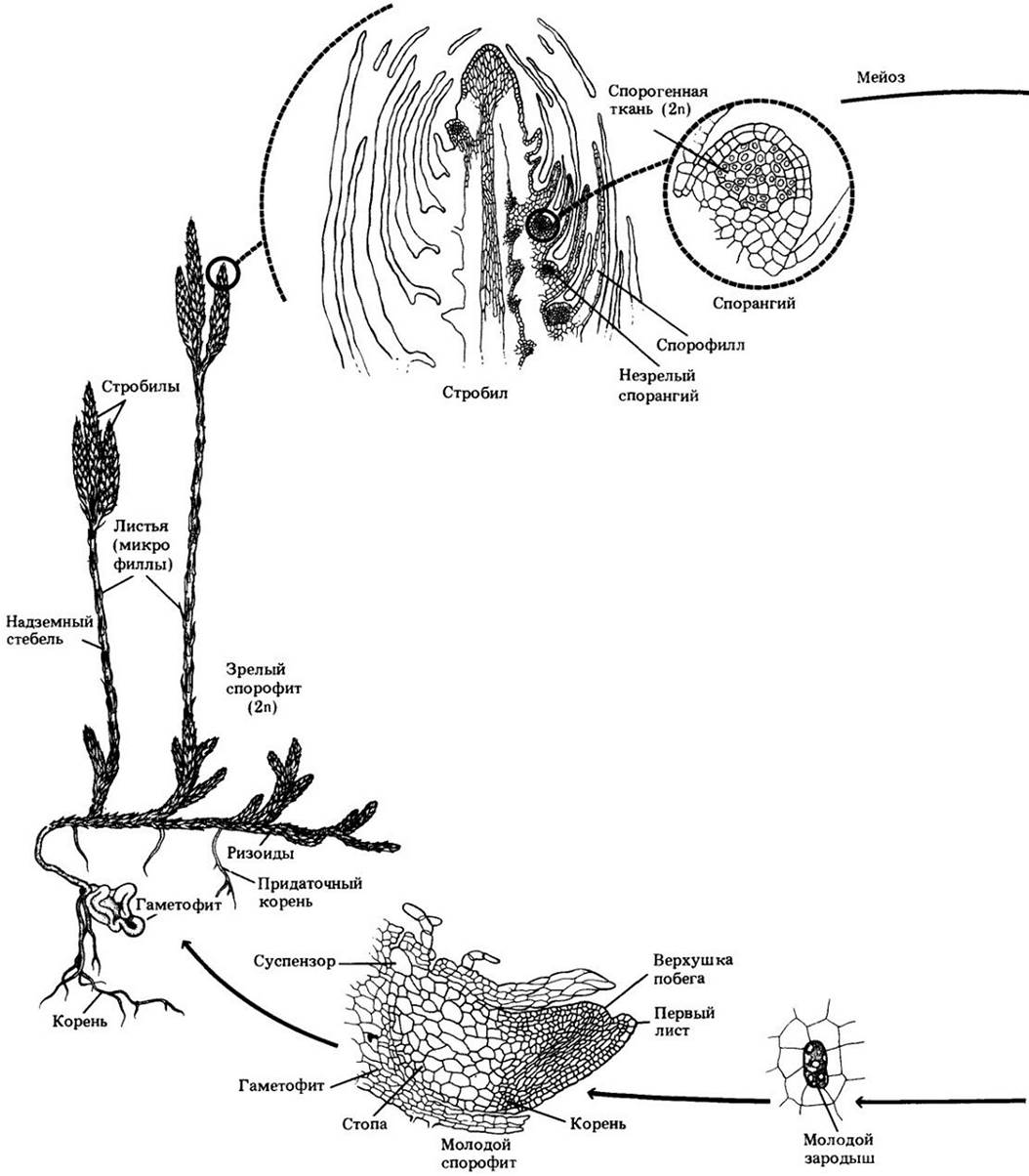
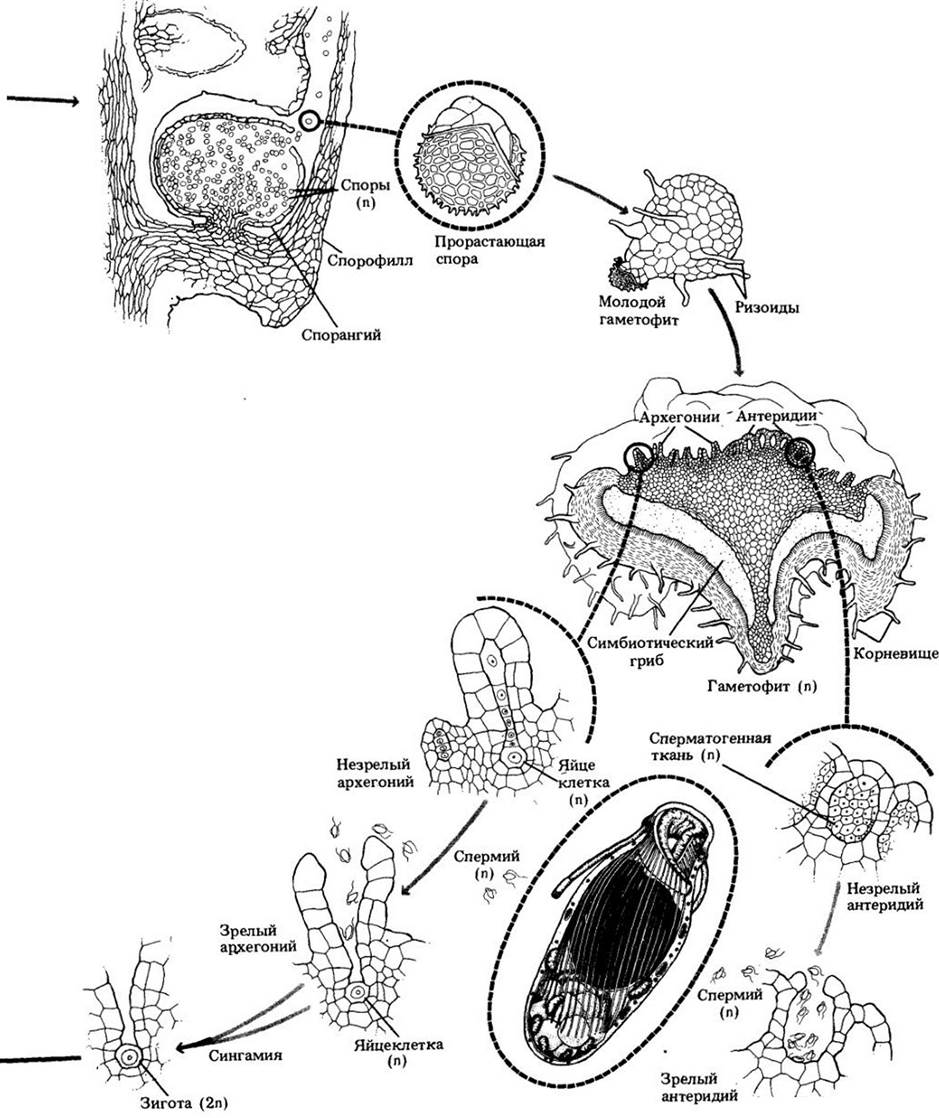
Рис. 17-18. Селагинелла. А. Поперечный срез стебля, на котором видны зрелые ткани. Протостела поддерживается в центре полого стебля удлиненными коровыми (эндодермальными) клетками, называемыми трабекулами. На рисунке видны только их фрагменты. Б. Протостела крупным планом
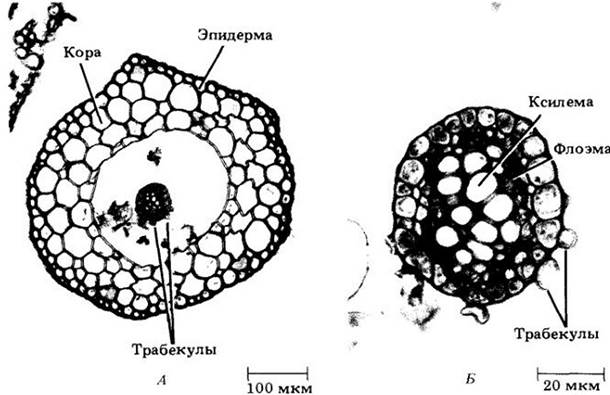
Рис. 17-19. Спорофит Isoetes muricata. Видны листья, похожие на иглы дикобраза, стебель и корни
Отдел Psilotophyta
Отдел включает два доживших до наших дней рода: псилот (Рsilotum) и тмезиптерис (Tmesipteris). Псилот распространен в тропиках и субтропиках; в США он встречается во Флориде, Луизиане, Аризоне, Техасе, на Гавайских островах и Пуэрто-Рико, а также как обычный оранжерейный сорняк. Ареал тмезиптериса ограничен Австралией, Новой Каледонией, Новой Зеландией и другими островами южной части Тихого океана. Оба рода — очень просто устроенные растения, напоминающие по своей основной структуре риниофиты.
Псилот уникален среди современных растений тем, что не имеет ни корней, ни листьев. Спорофит состоит из дихотомически ветвящейся надземной части с мелкими чешуевидными выростами и разветвленной подземной системы корневищ с многочисленными ризоидами (рис. 17-11). В наружных клетках коры корневищ присутствует эндомикоризный зигомицет. У псилота — протостела, следовательно, нет листовых прорывов (рис. 17-10).
Это растение равноспоровое; споры образуются в спорангиях, расположенных на концах коротких боковых ветвей. Прорастая, они дают начало обоеполым гаметофитам, которые похожи на фрагменты корневищ (рис. 17-11). Как и последние, подземный гаметофит содержит симбиотический гриб, а в некоторых случаях имеет проводящую ткань. Спермин псилота многожгутиковые и, чтобы достичь яйцеклетки, нуждаются в воде. Первоначально спорофит прикреплен к гаметофиту стопой — структурой, абсорбирующей питательные вещества из гаметофита. Впоследствии он от нее отделяется, а стопа остается погруженной в гаметофит.
Тмезиптерис растет как эпифит на древовидных папоротниках и других растениях (рис. 17-12). Листовидные придатки у него крупнее, чем чешуевидные выросты псилота, но в прочих отношениях эти два рода достаточно сходны.
Жизненный цикл псилота представлен на рис. 17-13.
Отдел Lycophyta
Четыре ныне живущих рода и примерно 1000 современных видов отдела Lycophyta (ликофиты, или плауновидные) представляют эволюционную линию, известную еще из девонского периода. Предками плауновидных почти с полной уверенностью можно считать зостерофиллофитов (см. рис. 17-8,5). Существует несколько порядков этого отдела; по крайней мере три из них, ныне вымершие, включали деревья, достигавшие иногда крупных размеров. Однако три современных порядка плауновидных состоят только из трав. И современные, и ископаемые ликофиты имеют микрофиллы; этот тип листьев — характернейшая черта отдела. Древовидные его представители были одними из доминирующих растений «углеобразующих» лесов каменноугольного периода (см. приложение к этой главе и рис. 18-1). Некоторые из них формировали структуры, сходные с семенами современных семенных растений. Большинство линий древесных плауновидных вымерло до конца палеозойской эры, т. е. не позже 248 млн. лет назад.
Lycopodium
Наверное, самыми известными из современных ликофитов являются плауны (род Lycopodium, см. рис. 10-7,5). Приблизительно 200 их видов встречаются от арктических областей до тропиков, хотя и редко играют заметную роль в растительных сообществах. Большинство тропических форм — эпифиты и поэтому редко бросаются в глаза, но несколько видов умеренных областей образуют ковры, иногда заметные в напочвенном покрове леса. Так как эти растения вечнозеленые, их лучше всего видно зимой.
Спорофит плауна состоит из ветвящегося корневища, от которого отходят надземные ветви и придаточные корни. И стебли, и корни протостелические (рис. 17-14). Микрофиллы обычно расположены спирально. Плаун — равноспоровое растение; спорангии расположены одиночно на верхней поверхности фертильных микрофиллов, называемых спорофиллами, — видоизменных листьев или листоподобных органов. У некоторых видов спорофиллы рассеяны среди стерильных микрофиллов и почти не отличаются от них (рис. 17-15). У других нефотосинтезирующие спорофиллы собраны в стробилы на верхушках надземных ветвей (см. рис. 10-7,5).
Споры плауна после прорастания дают начало обоеполым гаметофитам, которые в зависимости от вида представляют собой либо зеленые неправильно-лопастные образования, либо ветвящиеся подземные нефотосинтезирующие структуры. Как и у псилота и тмезиптериса, такие подземные гаметофиты вступают в симбиоз с грибом. Развитие и созревание архегониев и антеридиев у плаунов иногда продолжается от 6 до 15 лет. По мере роста гаметофита в последовательно созревающих архегониях может возникать целый ряд спорофитов.
Для оплодотворения необходима вода; двужгутиковый спермий подплывает в ней к архегонию и затем в ней же движется через его шейку. После оплодотворения зигота развивается в зародыш, растущий в брюшке архегония. Молодой спорофит может оставаться прикрепленным к гаметофиту в течение длительного времени, но в конце концов становится независимым.
Жизненный цикл плауна показан на рис. 17-17.
Selaginella
Среди современных родов плауновидных род селагинелла (Selaginella) насчитывает наибольшее число видов — около 700, распространенных главным образом в тропиках. Многие из них растут во влажных местообитаниях, но некоторые и в пустынных областях, впадая в покой вовремя наиболее сухой части года. Среди последних — так называемое «воскресающее» растение — Selaginella lepidophylla, ареал которого простирается от Мексики на север до Техаса и Нью- Мексико.
В основных чертах травянистые спорофиты селагинелл и плаунов схожи; здесь также наблюдаются микрофиллы, а спорофиллы образуют стробилы (рис. 17-16). В отличие от плаунов у селагинелл вблизи основания верхней поверхности каждого микрофилла и спорофилла развивается мелкий чешуевидный вырост, называемый язычком. Стебель и корень протостелические (рис. 17-18).
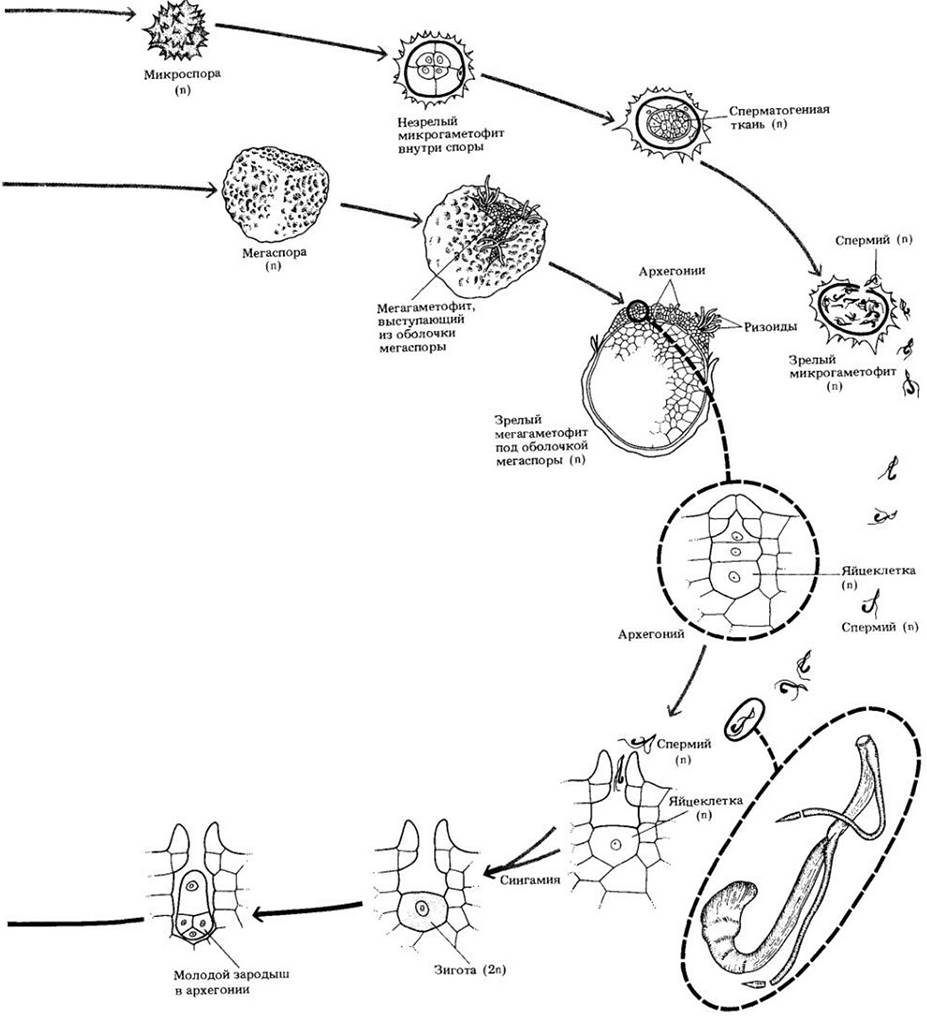
В отличие от равноспоровых плаунов селагинеллы разноспоровые и имеют однополые гаметофиты, в чем состоит наиболее существенное различие между этими двумя родами. Каждый спорофилл несет на своей верхней поверхности единственный спорангий. Мегаспорангии образуются на мегаспорофиллах, а микроспорангии — на микроспорофиллах. Оба типа спорангиев находятся в одном и том же стробиле.
Мужские гаметофиты (микрогаметофиты) у селагинелл возникают из микроспор. Четыре микроспоры образуются в результате мейоза из каждой материнской клетки микроспоры. Сам гаметофит развивается внутри микроспоры и лишен хлорофилла. В зрелом состоянии он состоит из единственной проталлиальной, или вегетативной, клетки и антеридия, образующего множество двужгутиковых спермиев. Для их высвобождения стенка микроспоры должна разорваться.
В ходе развития женского гаметофита (мегагаметофита) стенка мегаспоры разрывается, и та его часть, в которой формируются архегонии, выступает через разрыв наружу. Имеются данные, что в женском гаметофите иногда появляются хлоропласты, хотя более вероятно, что он получает питательные вещества в основном из запасов внутри мегаспоры.
Для того чтобы спермии достигли архегониев и произошло оплодотворение, требуется вода. Обычно перед этим гаметофиты отделяются от стробила. В ходе развития зародышей и у плаунов, и у селагинелл формируется структура, называемая суспензором (подвеском). У плаунов и некоторых видов селагинеллы он неактивен, но у других ее видов служит для «проталкивания» развивающегося зародыша вглубь богатой питательными веществами массы женского гаметофита. Растущий спорофит постепенно выступает из тканей гаметофита и становится независимым.
Жизненный цикл селагинеллы приведен на рис. 17-22.
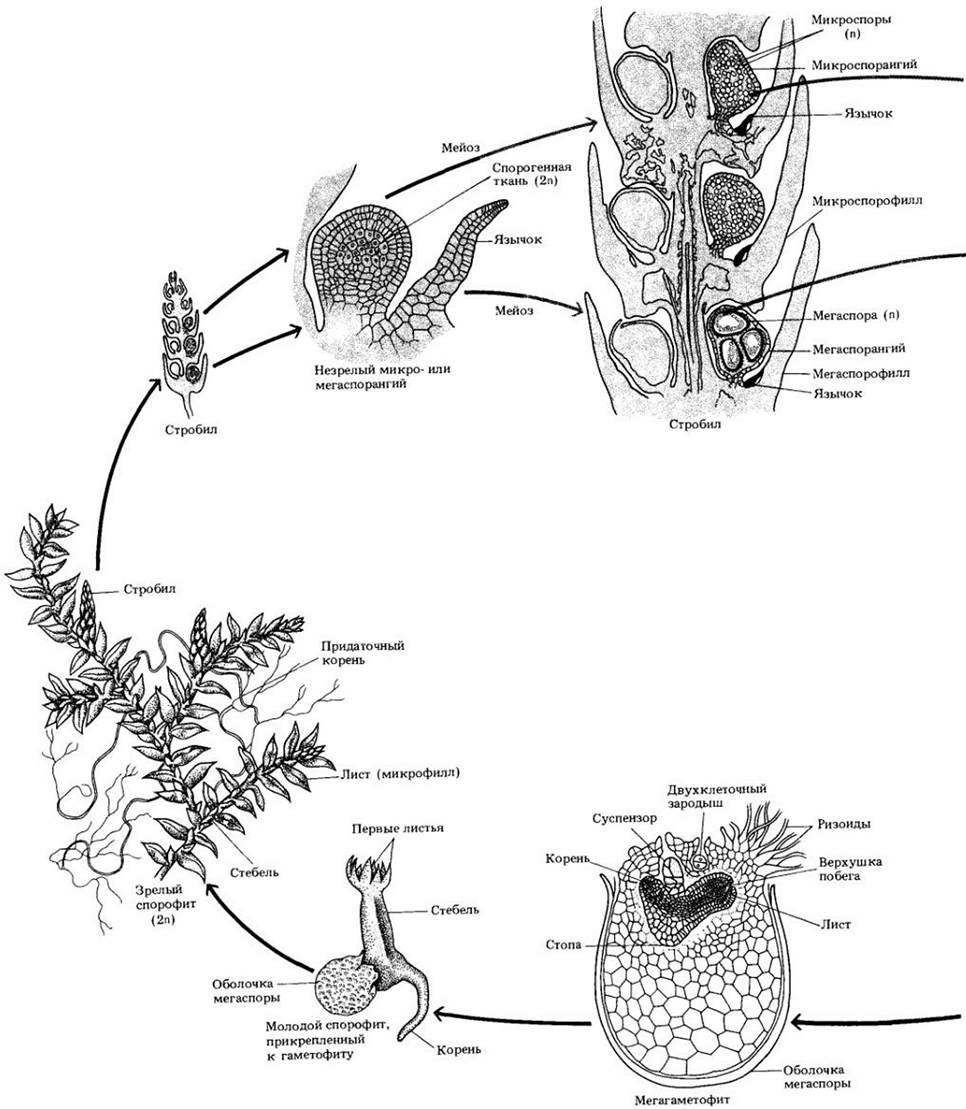
Рис. 17-22. Жизненный цикл селагинеллы — разноспорового растения. В одном и том же стробиле на спорофите образуются микро- и мегаспорангии. Микроспоры, формирующиеся в микроспорангиях, развиваются в мужские гаметофиты, а мегаспоры, возникающие в мегаспорангиях, — в женские гаметофиты. Микроспоры и мегаспоры падают на землю поблизости друг от друга, и спермиям нужно проплыть в воде до яйцеклетки совсем небольшое расстояние. Каждый спорангий располагается в пазухе чешуевидного придатка, язычка. У разноспоровых растений, подобных селагинелле, развитие гаметофитов начинается еще под оболочкой споры. Как и у семенных видов, молодой спорофит формируется среди тканей мегагаметофита, и главный источник питания для развивающегося зародыша — запасные вещества мегаспоры. Однако при этом нет периода покоя, характерного для многих семенных растений, а также интегументов, дающих начало семенной кожуре
Isoetes
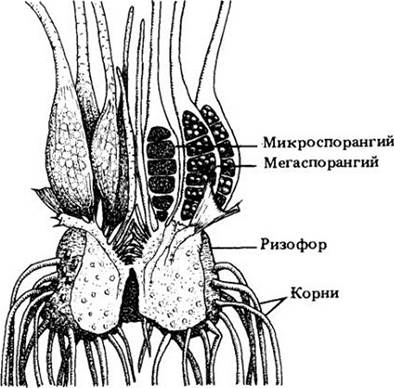
Замечательным представителем плауновидных является Isoetes, или полушник. Виды этого рода водные или растут в сезонно увлажняемых местах, пересыхающих в определенное время года. Спорофит полушника состоит из короткого мясистого подземного стебля (клубнелуковицы, или ризофора), несущего на верхней поверхности похожие на иглы дикобраза микрофиллы, а на нижней — корни (рис. 17-19). Каждый лист является потенциальным спорофиллом.
Как и селагинелла, полушник разноспоровый. Мегаспорангии образуются в основании мегаспорофиллов, а микроспорангии — микроспорофиллов, ближе к центру растения (рис. 17-20). Непосредственно над спорангием каждого спорофилла находится язычок.
Рис. 17-20. Схема вертикального разреза через полушник. Листья развиваются на верхней, а корни на нижней поверхности короткого мясистого подземною стебля. Одни листья (мегаспорофиллы) несут мегаспорангии, другие (микроспорофиллы) — микроспорангии. Микроспорофиллы располагаются ближе к центру растения
Одна из отличительных черт полушника — наличие специализированного камбия, образующего вторичные ткани клубнелуковицы. Наружу он откладывает только паренхиму, а внутрь — своеобразную проводящую ткань из ситовидных элементов, паренхимных клеток и трахеид, соотношение которых варьирует.
В 1984 г. было показано, что некоторые виды полушника из тропических высокогорий имеют уникальное свойство: они получают углерод для фотосинтеза не из атмосферы, а из осадочного материала. Листья этих растений лишены устьиц, покрыты толстой кутикулой и практически не осуществляют газообмена с атмосферой. У них фотосинтез типа САМ (см. с. 106), как и у некоторых видов полушника, засыхающих в определенное время года.
Отдел Sphenophyta
Подобно плауновидным, Sphenophyta (хвощевидные) известны еще из девонского периода, но наибольшего обилия и разнообразия они достигли в палеозое позднее, около 300 млн. лет назад. В позднем девоне и каменноугольном периоде они были представлены каламитами (см. с. 307) — деревьями, достигавшими в высоту 15 м, со стволами иногда более 20 см толщиной. В настоящее время хвощевидные представлены единственным травянистым родом Equisetum (хвощ), включающим 15 видов (рис. 17-21).
Рис. 17-21. Один из видов хвоща с практически лишенными хлорофилла фертильными побегами, резко отличающимися по внешнему облику от вегетативных. А. Фертильные побеги, каждый с верхушечным стробилом; на каждом узле мутовки чешуевидных листьев. Б. Вегетативные побеги
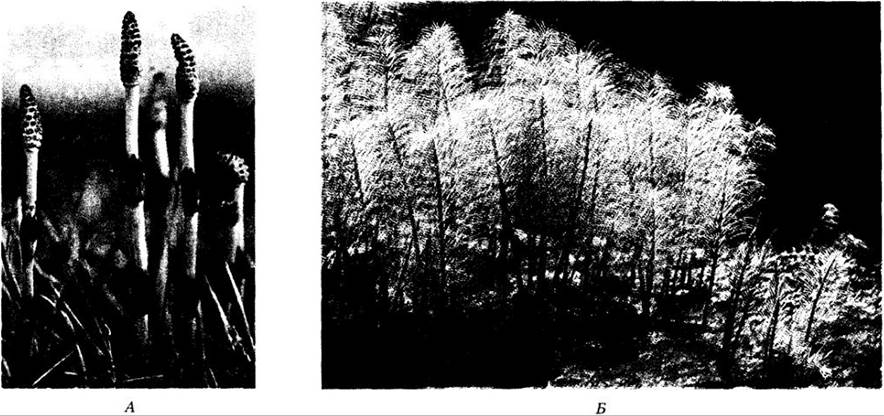
Виды хвоща широко распространены во влажных или болотистых местах, около рек и вдоль лесных опушек (рис. 17-21). Их легко узнать по явно членистым стеблям и грубой текстуре. Мелкие чешуевидные листья, простые по строению, но, вероятно, являющиеся редуцированными мегафиллами, собраны в узлах мутовками. Ветви, если они имеются, отходят от узлов латерально, чередуясь с листьями. Междоузлия (части стеблей между соседними узлами) имеют ребра (гребни), жесткие из-за отложения кремнезема в эпидермальных клетках. Из-за своей жесткости хвощи использовались для чистки посуды, особенно в эпоху колонизации Америки европейцами, и получили в то время название «камышовых мочалок». Корни придаточные, отходят от узлов корневищ.
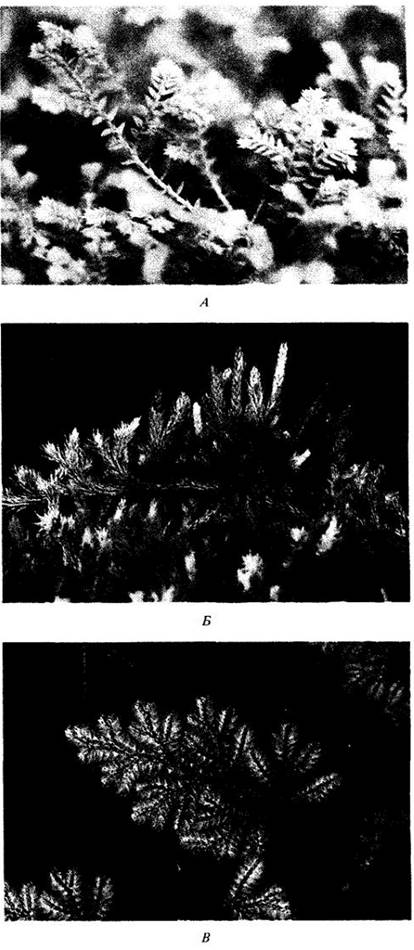
Рис. 17-25. Разнообразие папоротников. А. Osmunda cinnamomea. Б. Древовидный папоротник Dicksonia squarrosa из Новой Зеландии. В. Растущий на известняке вид Notholaenaneglecta, встречающийся в шт. Техас, Аризона, Нью-Мексико (США). Г. Разноспоровый водный папоротник Marsilea

Надземные побеги хвощей, иногда отмирающие в неблагоприятные сезоны, образуются на многолетних разветвленных корневищах. Надземный стебель сложен по анатомическому строению (рис. 17-23). В зрелом состоянии полая «сердцевина» междоузлий окружена кольцом более мелких, так называемых каринальных каналов, каждый из которых связан с тяжем первичной ксилемы и первичной флоэмы.
Рис. 17-23. Анатомия стебля хвоща. А. Поперечный срез, на котором видны зрелые ткани. Б. Отдельный проводящий тяж с ксилемой и флоэмой
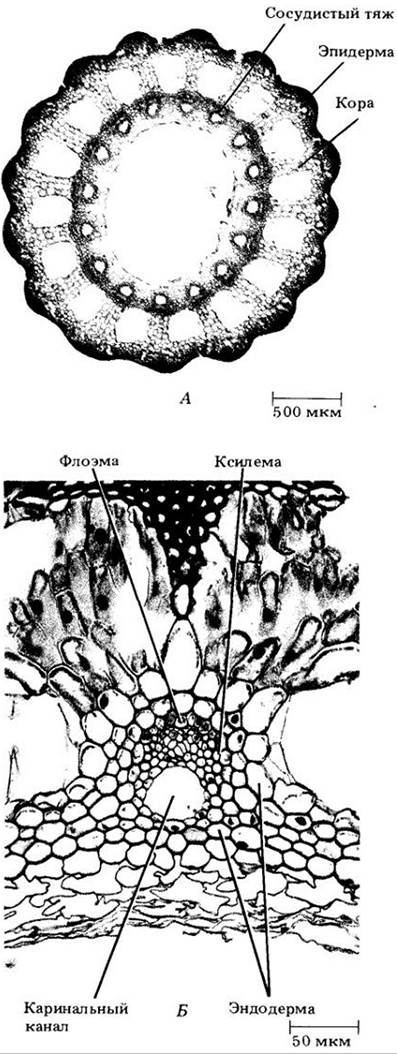
Рис. 17-26. Жизненный цикл хвоща. В общих чертах он идентичен жизненным циклам псилота, плауна и равноспоровых папоротников. Мейоз происходит в спорангиях, расположенных по краю зонтиковидных, собранных в стробилы структур, спорангиофоров. При созревании спор элатеры, прикрепленные к их оболочке, вероятно, способствуют их рассеиванию из раскрывающегося спорангия. Гаметофиты зеленые, свободноживущие, обоеполые (как на этом рисунке) или мужские. Для достижения яйцеклетки многожгутиковым спермиям требуется вода. У хвощей, как и у других споровых сосудистых растений, развитие зародыша происходит в брюшке архегония, а молодой спорофит прикрепляется к гаметофиту стопой. В конце концов спорофит отделяется от нее и становится независимым организмом. У хвощей в отличие от прочих споровых сосудистых растений он имеет членистые, ребристые стебли.
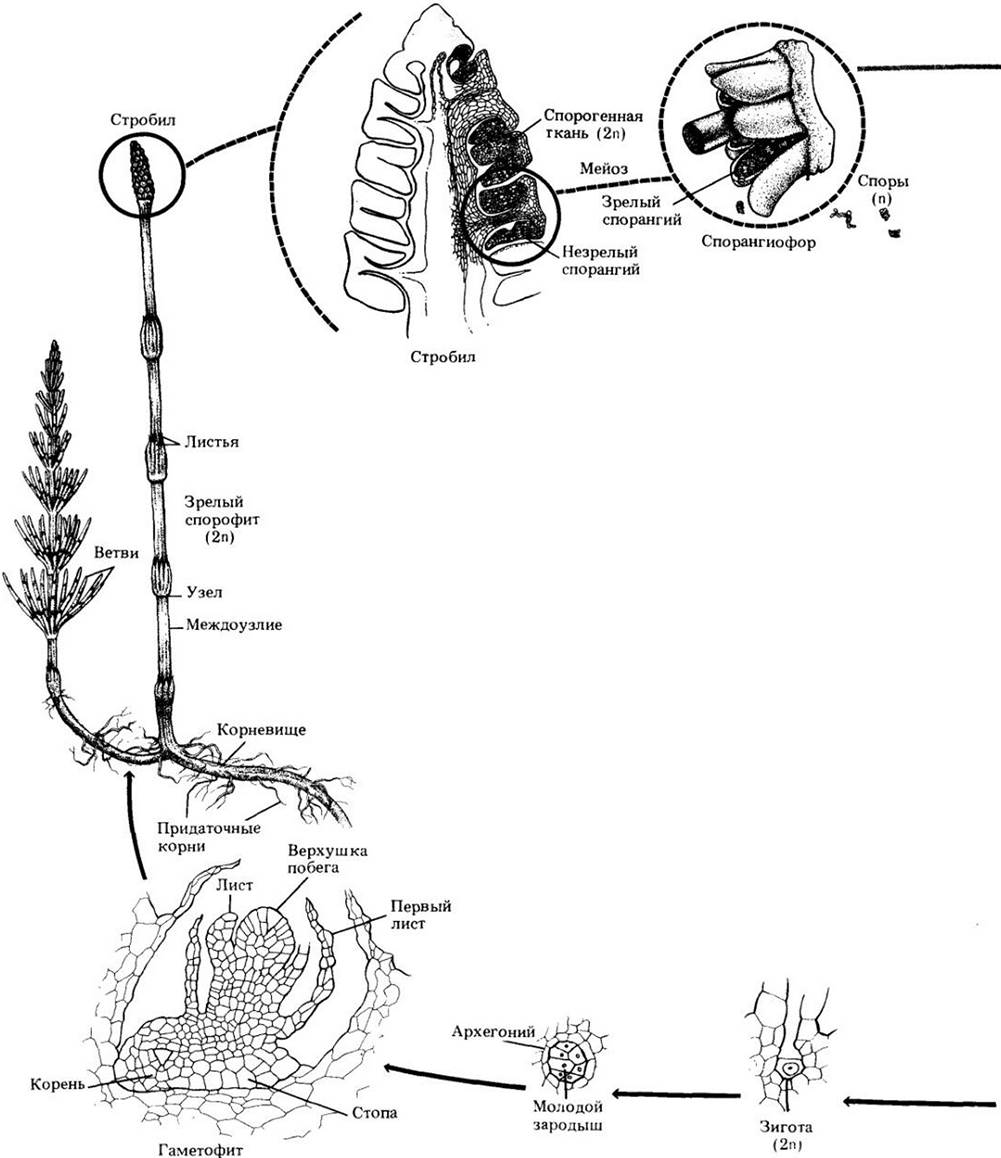
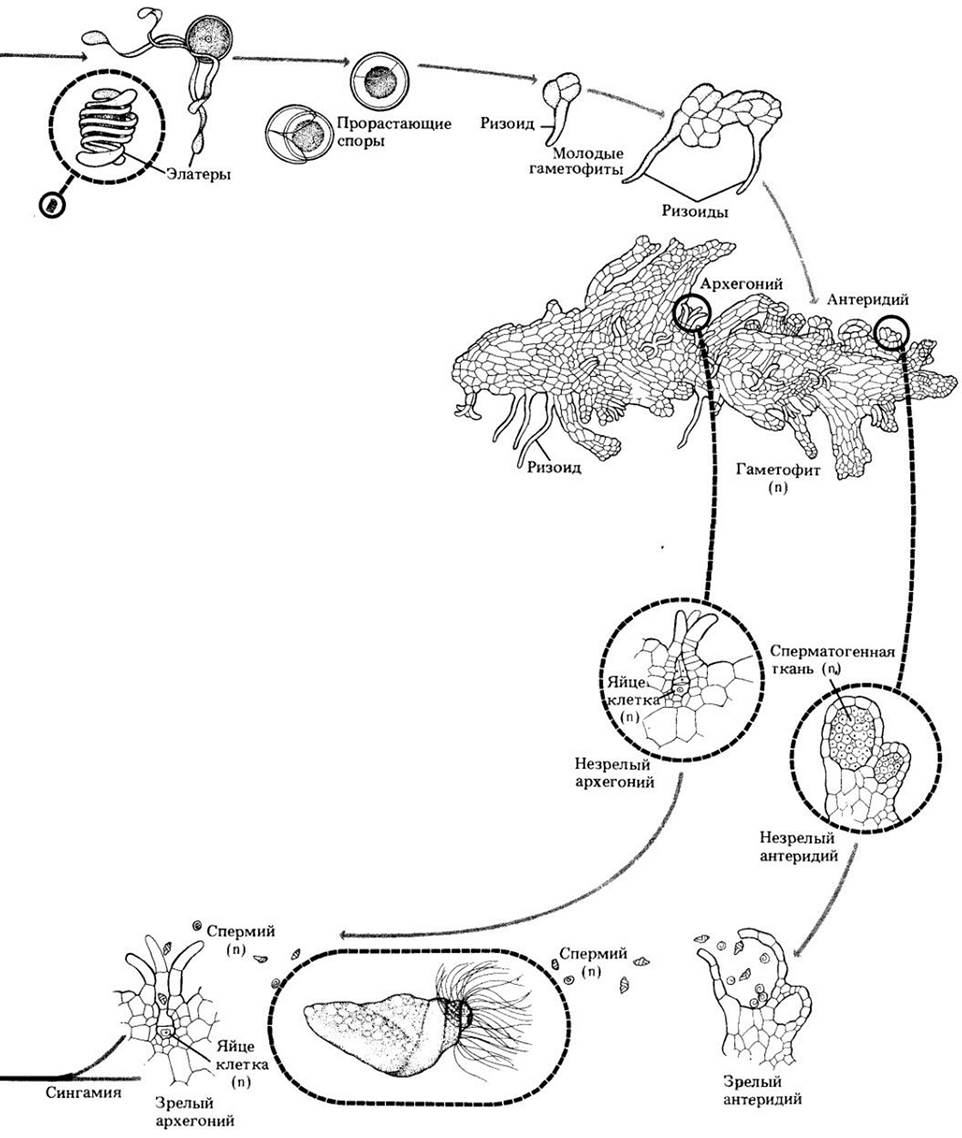
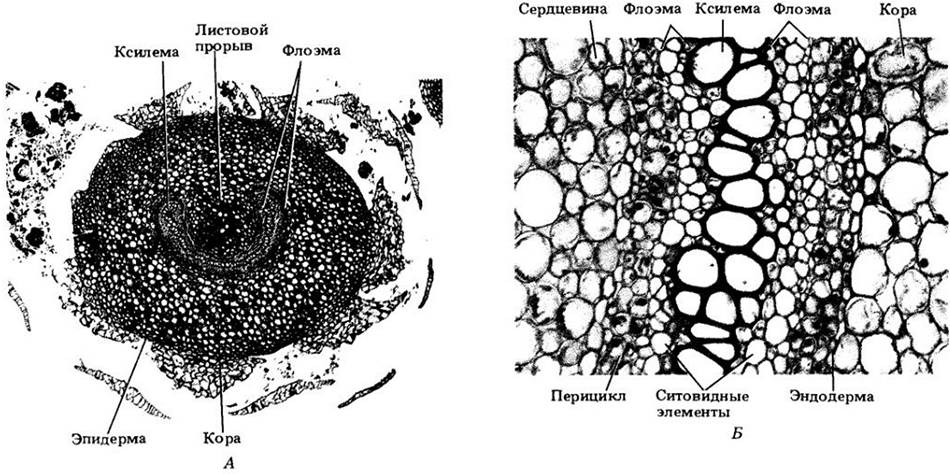
Рис. 17-27. Анатомическое строение корневищ папоротников. A. Adiantum. Поперечный срез корневища; видна сифоностела. Обратите внимание на широкий листовой прорыв. Б. Поперечный разрез через проводящую зону корневища древовидного папоротника Dicksonia. Флоэма состоит главным образом из ситовидных элементов, а ксилема только из трахеид
Хвощи равноспоровые растения. Спорангии образуются группами по пять-десять вдоль края мелких зонтиковидных структур, спорангиофоров (спорангиеносных веточек), собранных в стробил на верхушке стебля (см. рис. 17-21, A и 17-26). Фертильные стебли некоторых видов содержат мало хлорофилла, резко отличаются от вегетативных стеблей и часто появляются раньше них в начале весны (см. рис. 17-21). У других видов стробилы образуются на концах вегетативных стеблей (см. рис. 10-7,0- Когда многочисленные споры созревают, спорангии сжимаются и расщепляются вдоль внутренней поверхности, высвобождая их. Элатеры, отходящие от наружного слоя оболочки споры, свернуты спиралью во влажном состоянии и разворачиваются в сухом, что, как полагают, играет определенную роль в рассеивании спор.
Гаметофиты хвощей зеленые, свободноживущие, большинство из них размером с булавочную головку. Они развиваются главным образом на иле, незадолго перед этим заливавшемся водой и потому богатом питательными веществами. Гаметофиты (рис. 17-24) достигают половой зрелости через 3 — 5 недель и являются либо обоеполыми, либо мужскими. На обоеполых архегонии развиваются раньше антеридиев, что увеличивает вероятность перекрестного оплодотворения. Спермин много жгутиковые; для достижения яйцеклеток им требуется вода. На одном гаметофите могут быть оплодотворены сразу несколько яйцеклеток, из которых затем развиваются зародыши, т. е. молодые спорофиты.
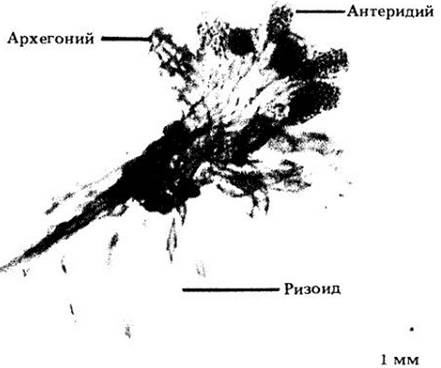
Рис. 17-24. Обоеполый гаметофит хвоща с мужскими и женскими гаметангиями. Сравните его с изображенным на рис. 16-2 гаметофитом из Шотландии, возраст которого 400 млн. лет
Жизненный цикл хвощей представлен на рис. 17-26.
Рис. 17-28. Улиткообразно свернутые листья Osmunda cinnamomea. Раньше их употребляли в пищу, однако сейчас имеется серьезное подозрение, что они в какой-то мере ядовиты

Рис. 17-29. Сорусы — скопления спорангиев на нижней поверхности листьев папоротников. А. У Dennstaedtia punciilobula и других видов этого рода сорусы голые. Б. У орляка (Pteridium aquilinum), представленного на рисунке, а также у Adiantum сорусы расположены вдоль краев листовой пластинки, которые загибаются, прикрывая их. В. У вечнозеленого лесного папоротника Dryopteris marginalis сорусы, также находящиеся у краев листовых пластинок, полностью покрыты почковидными индузиями. Г. У Onoclea sensibilis спорангии расположены внутри шаровидных долей особых, сильно отличающихся от стерильных листьев. Стенки спорангиев многих папоротников состоят только из одного слоя клеток и имеют специализированные механизмы растрескивания. Однако у некоторых примитивных групп этого отдела их стенки многослойные и разрываются по единственной линии клеток с тонкими оболочками
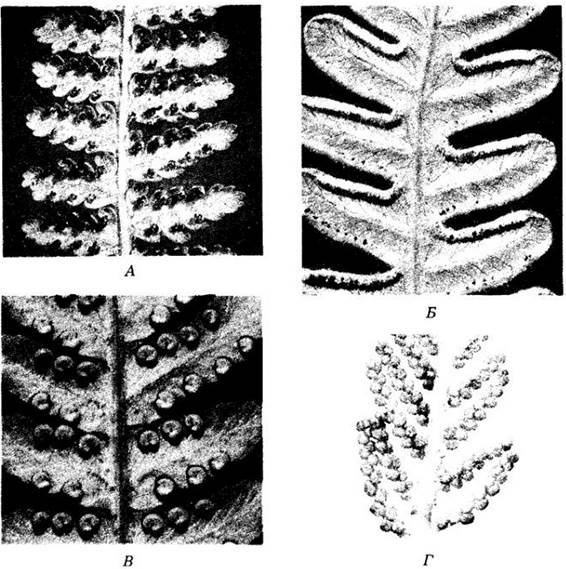
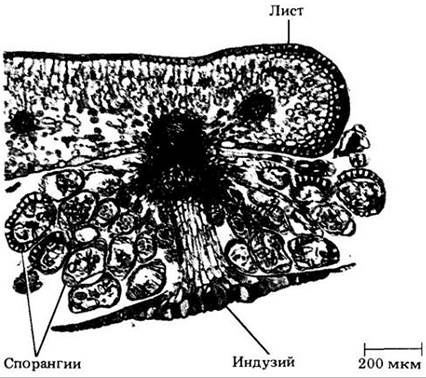
Рис. 17-30. Равноспоровый папоротник Cyrtomium falcatum. Поперечный срез листа с сорусом на нижней поверхности. Находящиеся на разных стадиях развития спорангии прикрыты зонтиковидным индузием
ОтделТ Рterophyta
Представители этого отдела (папоротниковидные) относительно обильно представлены ископаемыми остатками, начиная с каменноугольного периода (см. приложение к этой главе и рис. 18-1), и дожили до настоящего времени, а некоторые их группы встречались даже в девонском периоде. Около двух третей из приблизительно 12 000 современных видов растут в тропиках, а оставшаяся треть населяет умеренные области земного шара, включая пустынные районы. Папоротниковидные намного превосходят по числу видов все прочие группы современных споровых сосудистых растений.
Представители этого отдела весьма разнообразны по жизненным формам и занимаемым местообитаниям (рис. 17-25). Некоторые из них сильно отличаются от привычных нам папоротников по внешнему виду; например, Salvinia, водное растение с цельными листьями до 2 см длиной. Более «папоротникоподобными» являются плавающие водные виды рода Azolla (см. рис. 26-16), играющие важную роль в сельскохозяйственных и природных экосистемах, так как в полостях при основании их листьев поселяются симбиотические азотфиксирующие цианобактерии. Другая крайность — древовидные папоротники (рис. 17-25, Б), например, из рода Cyathea, среди которых зарегистрированы растения более 24 м высотой с листьями длиной более 5 м. Хотя их стволы могут достигать более 30 см в диаметре, ткани здесь полностью первичные по происхождению. Камбий известен только у гроздовника (Botrychium), относительно мелкого травянистого растения.
Большинство лесных папоротников умеренных областей имеют мясистые, сифоностелические корневища (рис. 17-27), образующие каждый год новые пучки листьев. Корни придаточные, отходят от корневищ у оснований листьев. Листья, или вайи, являются мегафиллами и составляют наиболее заметную часть спорофита. Соотношение их поверхности и объема показывает, что это намного более эффективные фотосинтезирующие органы, чем микрофиллы плауновидных. Папоротники — единственные споровые сосудистые растения с мегафиллами. Вайи обычно сложные, т. е. их пластинка разделена на листочки, прикрепленные к стержню, рахису, продолжающему ножку листа, или черешок. Почти у всех папоротников молодые листья в почке улиткообразно закручены (рис. 17-28), т. е. наблюдается так называемое свернутое листосложение. Оно обусловлено тем, что нижняя поверхность вайи на ранней стадии развития растет быстрее, чем верхняя. Это регулируется гормоном ауксином, продуцируемым молодыми листочками на внутренней стороне «улитки».
За исключением нескольких родов, все папоротники равноспоровые. Спорангии располагаются различными способами на нижней поверхности обычных листьев, на видоизмененных вайях или на особых осях (рис. 17-29). Они обычно собраны в кучки, называемые сорусами. У многих родов сорусы покрыты особыми выростами листа — покрывальцами, или индузиями, которые могут сморщиваться при созревании спорангиев (рис. 17-30). В это время зрелые споры, образовавшиеся из материнских клеток спор в результате мейоза, выбрасываются наружу благодаря резкому разрыву клеток в участке спорангия, называемом стомием. Спорангии имеют ножки, и в оболочке каждого из них находится особый слой клеток с неравномерно утолщенной стенкой, кольцо (аннулюс). Сжатие кольца вызывает разрыв клеток стомия, а следующее затем его внезапное распрямление — выстреливание спор, напоминающее катапультирование.
Разноспоровость среди современных папоротников известна только у двух специализированных водных групп (см. рис. 17-25, Г); некоторые вымершие папоротниковидные также были разноспоровыми.
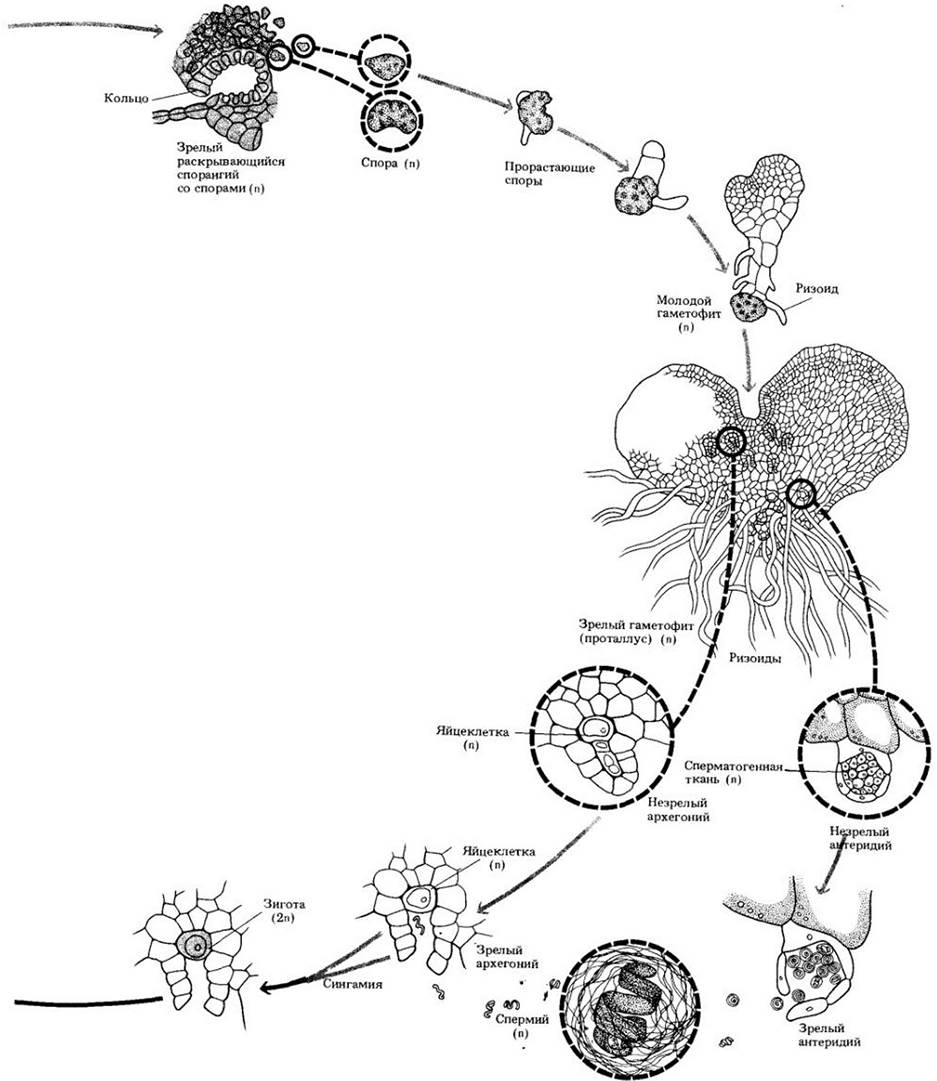
Споры большинства равноспоровых папоротников дают начало свободноживущим обоеполым гаметофитам. Гаметофит начинает свое развитие с крошечной бледно-зеленой, похожей на водоросль цепочки клеток, называемой протонемой. Затем из нее образуется плоская сердцевидная пленчатая структура, проталлус (заросток), с многочисленными ризоидами в центре нижней поверхности. Антеридии и архегонии также формируются на его нижней поверхности. Антеридии обычно появляются раньше, главным образом среди ризоидов, а архегонии позднее, вблизи выемки на переднем конце гаметофита. Разница во времени формирования двух типов гаметангиев способствует перекрестному оплодотворению папоротников. Как равноспоровым, так и разноспоровым представителям отдела для передвижения многожгутиковых спермиев к яйцеклеткам требуется вода.
На ранней стадии своего развития зародыш, т. е. молодой спорофит, получает питательные вещества от гаметофита через стопу. Однако он растет очень быстро и вскоре становится независимым организмом. К этому времени гаметофит отмирает.
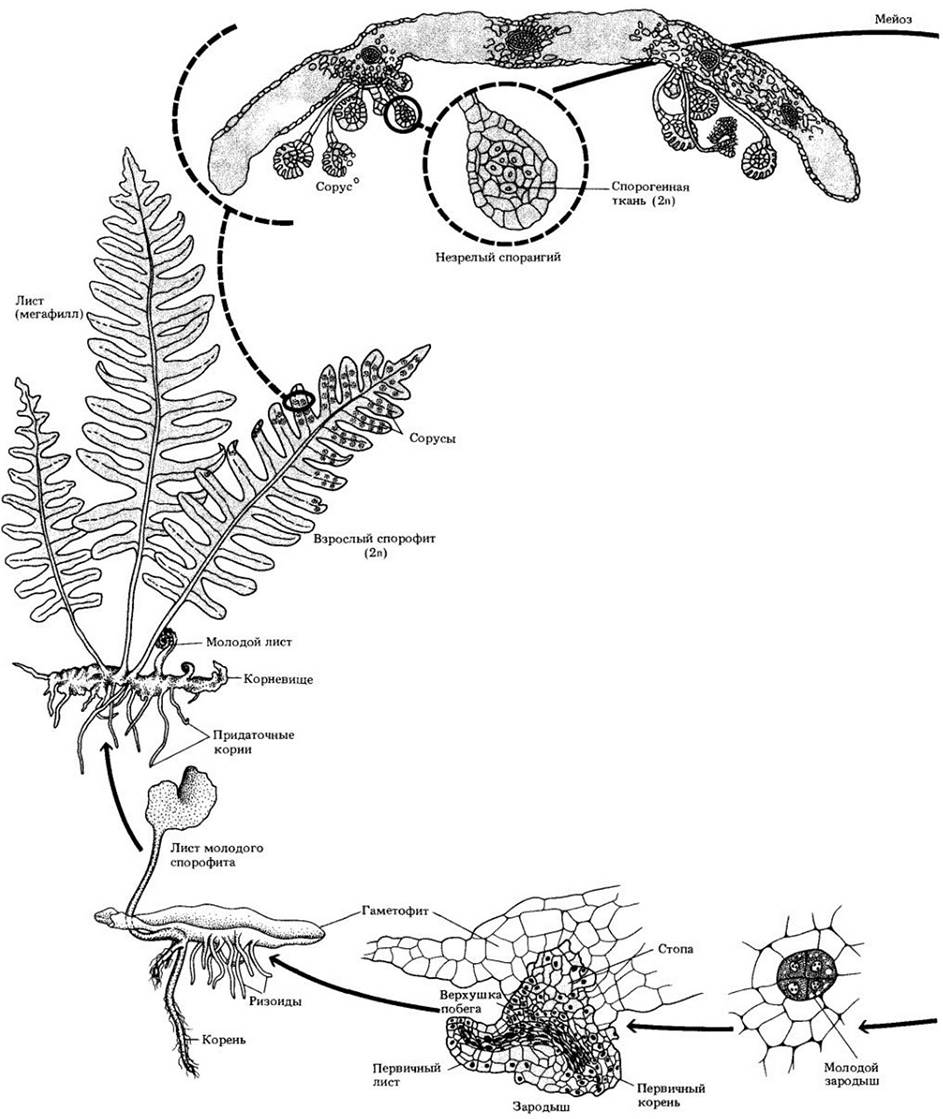
Жизненный цикл равноспорового папоротника показан на рис. 17-31.
Рис. 17-31. Жизненный цикл равноспорового папоротника Polypodium. После мейоза в спорангиях образуются споры, дающие после рассеивания гаметофиты, которые у большинства видов зеленые и независимые в своем питании. Многие из них толщиной всего в один слой клеток, более или менее сердцевидные, с апикальной выемкой; другие толще и могут быть неправильной формы. От нижней поверхности гаметофита отходят, проникая в субстрат, специализированные клеточные нити (ризоиды).
На нижней поверхности гаметофита находятся колбовидные архегонии с погруженной в его ткань расширенной нижней частью. Их шейки состоят из нескольких ярусов клеток. Антеридии также находятся на нижней поверхности гаметофита и окружены стерильным покровным слоем. В них образуются многочисленные, спирально закрученные многожгутиковые спермии. Когда они созревают, то при достаточном увлажнении антеридии лопаются, высвобождая спермии, заплывающие в шейку архегония. В его брюшке происходит оплодотворение, и возникшая зигота сразу же начинает делиться. Молодой зародыш растет и дифференцируется непосредственно во взрослый спорофит, некоторое время получая питание от гаметофита, но вскоре начиная достаточно интенсивно фотосинтезировать для поддержания своего независимого существования. После укоренения молодого спорофита в почве гаметофит разрушается