СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 18. СЕМЕННЫЕ РАСТЕНИЯ
Одним из важнейших новоприобретений в эволюции сосудистых растений было семя. Оно является, вероятно, одним из факторов, определивших господство семенных растений в современной флоре — господство, постепенно усилившееся на протяжении нескольких миллионов лет (рис. 18-1). Причина этого проста: семя способствует выживанию.
Рис. 18-1. Реконструкция верхнекарбонового заболоченного леса с господством древовидных ликофитов лепидодендронов (образуют верхний ярус древостоя в левой части рисунка). В молодости они не разветвленные (напоминают ершики для мытья бутылок), затем слабо и наконец сильно ветвятся, создавая лесной полог. На возвышенном участке (в правой части рисунка) видна сигиллярия (Sigillaria) — ликофит, приспособленный к более сухим условиям (дерево с пучками длинных листьев). Каламиты (гигантские хвощи, изображенные с левого края и на переднем плане в центре) и древовидные папоротники рода Psaronius (со стволами, суживающимися кверху; с правого края) — также споровые сосудистые растения. Довольно много и семенных растений, среди которых медуллоза (Medullosa), семенной папоротник с вилъчато разделенными вайями (в центре и с правого края) и кордаиты, представленные как мангровидными формами с погруженными в воду корнями (справа внизу), так и высокими деревьями (справа в верхнем ярусе)


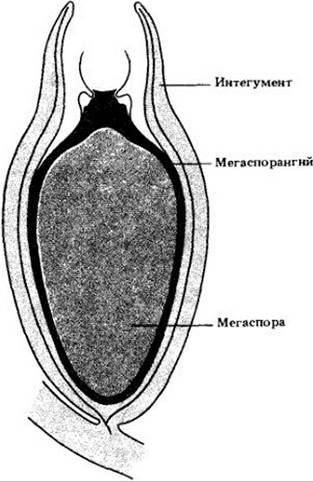
Все семенные растения разноспоровые, и их крайне редуцированный мегагаметофит заключен внутри мегаспоры. Та в свою очередь находится в мясистом мегаспорангии, называемом здесь нуцеллусом. В отличие от бессемейных разноспоровых растений он покрыт одним или двумя дополнительными слоями ткани, интегументами. Они полностью заключают в себя мегаспорангий, оставляя лишь отверстие (микропиле) на его верхушке. Нуцеллус и интегумент(ы) вместе называются семязачатком (рис. 18-2).
Рис. 18-2. Разрез семязачатка Eurystoma angulare, показывающий взаимное расположение интегумента, мегаспорангия (нуцеллуса) и мегаспоры. Оплодотворение приводит к превращению семязачатков в семена; другими словами, семена — оплодотворенные зрелые семязачатки
После оплодотворения интегументы превращаются в семенную кожуру, и формируется семя. Другими словами, оно развивается из семязачатка, и его часто называют зрелым семязачатком. У большинства современных семенных растений зародыш, или молодой спорофит, формируется внутри семени до его опадания, однако у многих древних представителей этой группы (см., например, рис. 18-2 — 18-4) семена опадали, вероятно, до развития зародыша. Возможно, первый способ обеспечивает лучшее выживание в холодных и суровых условиях, а пермский период, когда возникли хвойные, саговниковые и гинкговые, несомненно, был временем климатических крайностей. Кроме зародыша и кожуры, у всех семян имеется запас питательных веществ, что еще больше увеличивает шансы молодого растения на выживание.
Древнейшие из известных семян относятся к позднему девону (около 360 млн. лет назад) (рис. 18-3). В следующие 50 млн. лет появилось множество семенных растений, в частности семенные папоротники, кордаитовые и хвойные (рис. 18-4). Первые сосудистые растения, включая три вымерших отдела, описанных в гл. 17, были по сравнению со своими семенными потомками распространены очень широко. Объясняется это тем, что семена не рассеиваются на такие большие расстояния, как споры. С появлением семенных форм флоры, т. е. совокупности видов в определенных областях планеты, стали сильнее отличаться друг от друга.
Рис. 18-3. А. Реконструкция фертильной ветви позднедевонского растения Archeosperma arnoldis с семяподобными структурами. Купулы — бокаловидные образования, частично заключавшие в себе мегаспорангии, располагались парами; в каждой купуле находились два колбовидных семени длиной около 4 мм с разделенными на лопасти верхушками. Б. Схема расположения мегаспоры в семени. В. Мегаспора, извлеченная из семени путем мацерации. Эта ископаемая находка из Пенсильвании — древнейшее из известных семян (возраст 360 млн. лет)
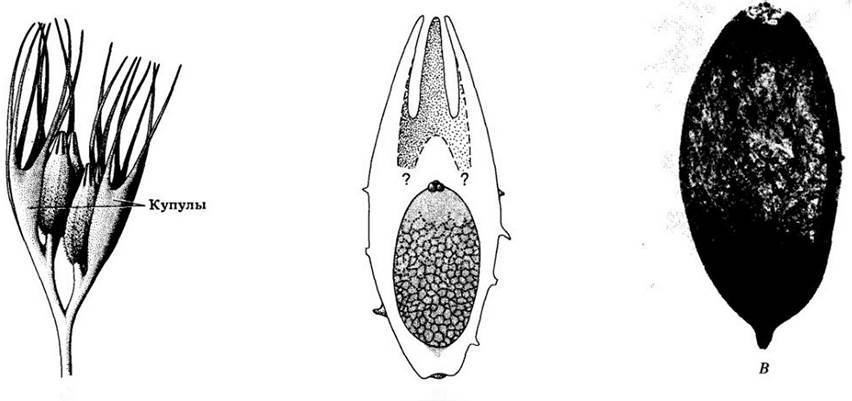
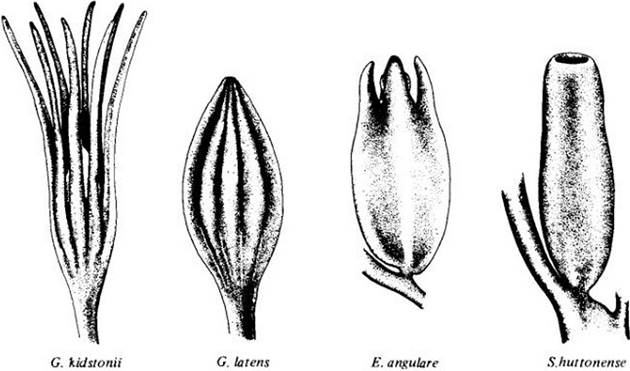
Рис. 18-4. Семяподобные структуры нескольких палеозойских растений, позволяющие проследить эволюцию семян. У Genomosperma kidstonii (genomein по-гречески «становиться». а sperma — «семя») нуцеллус был окружен 8 — 11 отдельными пальцевидными выро- cma.au, отходящими от основания мегаспорангия. Интегументов еще не было. У Genomosperma laiens интегументарные лопасти срослись примерно на треть длины, образуя простой покров семени. У Eurystoma angulare слияние почти полное, а у Siamnostoma huitonense оно уже завершилось
Семенные растения (все они имеют мегафиллы) включают пять отделов с ныне живущими представителями: саговниковые (Суcadophyta), гинкговые (Ginkgophyta), хвойные (Coniferophyta), гнетовые (Gnetophyta) и цветковые (Anthophyta). Прежде чем рассмотреть их, мы кратко остановимся на еще одной группе бессемейных сосудистых — прогимноспермах, известных только в ископаемом состоянии. Они обсуждаются здесь, а не в гл. 17, так как являются вероятными предками голосеменных.
Покрытосеменные, или цветковые, в настоящее время господствующая группа сосудистых растений, достоверно известны начиная с раннего мела (примерно 127 млн. лет назад). Их эволюция рассматривается и сравнивается с эволюцией голосеменных в гл. 29. Хотя покрытосеменные должны быть несколько древнее, чем дошедшие до нас их ископаемые остатки, все же в ходе разносторонней эволюции сосудистых растений они возникли сравнительно недавно.
Прогимноспермы
В позднем палеозое существовала группа растений, называемых прогимноспермами, промежуточная по своим признакам между тримерофитами и голосеменными. Хотя прогимноспермы размножались свободно рассеиваемыми спорами, у них была вторичная ксилема, очень сходная с присутствующей у голосеменных (рис. 18-5). От всех девонских деревьев они отличались также образованием вторичной флоэмы. Прогимноспермы и палеозойские папоротники, вероятно, являются потомками более древних тримерофитов (см. рис. 17-8, В), от которых они отличались главным образом более сложным ветвлением и соответственно более развитыми проводящими системами. Возможно также, что папоротники произошли от прогимноспермов. Так как тримерофиты дали начало последним, а папоротники возникли от одной из этих групп, вопрос только во времени появления папоротников.
Рис. 18-5. Радиальный разрез вторичной ксилемы, или древесины, представителя прогимноспермов Callixylon newberryi. Эта ископаемая древесина с правильными рядами трахеид с порами удивительно похожа на древесину некоторых голосеменных

Наиболее важное прогрессивное отличие прогимноспермов от тримерофитов — присутствие бифациального камбия, т. е. камбия, образующего вторичные ксилему и флоэму. Такой его тип характерен для семенных и, по-видимому, впервые появился именно у прогимноспермов.
Некоторые из представителей этого отдела (группа аневрофитона), жившие в девонский период (примерно 360 — 380 млн. лет назад), характеризовались трехмерным ветвлением (рис. 18-6) и имели протостелы. По своему строению они были похожи на некоторые древние семенные папоротники, что дало некоторым палеоботаникам основание считать системы ветвей этих растений возможными предшественниками вайеподобных листьев последних.
Рис. 18-6. Реконструкция ветвления Triloboxylon ashlandicum, представителя прогимноспермов ив группы аневрофитона (Aneurophyton). Главная ось несет вегетативные ветви (на рисунке вверху и внизу), а между ними системы осей со спорангиями
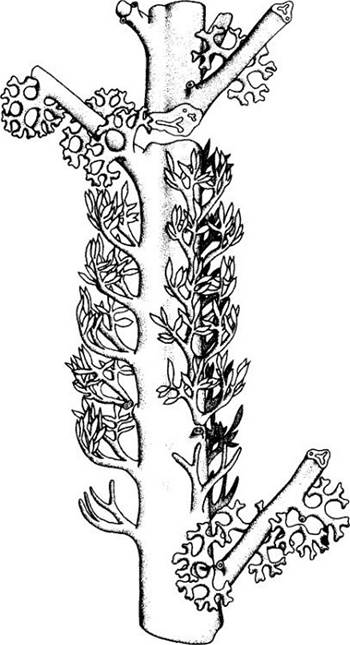
Другая крупная группа прогимноспермов — археоптерисовые — также появилась в девоне (около 370 млн. лет назад) и вымерла в раннем карбоне, около 340 млн. лет назад (рис. 18-7). Она считается более продвинутой, так как латеральные системы ветвей здесь были сплюснуты в одной плоскости и несли пластиновидные структуры, своего рода листья (рис. 18-8). Эти облиственные ветви сходны у них с побегами древних хвойных. Более крупные ветви археоптерисовых имели сердцевину. Большинство прогимноспермов были равноспоровыми, но у некоторых видов Archaeopteris обнаружена разноспоровость.
Рис. 18-7. Реконструкция археоптериса из прогимноспермов, ископаемые остатки которого часто встречаются на востоке Северной Америки. Виды этого рода достигали в высоту более 20 м, и некоторые из них, по-видимому, образовывали целые леса

Рис. 18-8. Реконструкция вайеподобной системы боковых ветвей Archaeopteris macilenta из прогимноспермов. На ветвях первого порядка в средней части видны фертильные листья с созревающими спорангиями

Ископаемые стволы археоптериса (Аrchaeopteris) достигают более метра в диаметре и до 10 м в длину, т. е. по крайней мере некоторые виды этой группы были крупными деревьями. Пространственное распределение находок наводит на мысль о доминировании археоптериса в обширных лесах некоторых областей. Как видно по реконструкции на рис. 18-7, своим ветвлением он мог напоминать хвойные, однако это только предположение.
Морфологические данные, накопившиеся за несколько последних десятилетий, вполне подтверждают мнение об эволюции голосеменных от прогимноспермов по мере развития семени. Однако невозможно с уверенностью сказать, возникало оно в этой эволюционной линии один раз или несколько. Не ясно также, от какой группы (или групп) прогимноспермов произошли голосеменные.