СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ VIII. ЭВОЛЮЦИЯ
ГЛАВА 29. ЭВОЛЮЦИЯ ЦВЕТКОВЫХ РАСТЕНИЙ
Среди современных растений значительное место принадлежит покрытосеменным. Деревья и кустарники, луга и сады, пшеничные и кукурузные поля, полевые цветы, овощи и фрукты на прилавках, красочные композиции в витринах цветочных магазинов, герань на камине, ряска и кувшинки в пруду, взморник на заливе, кактусы в пустыне — где бы мы ни находились, везде встретимся с ними.
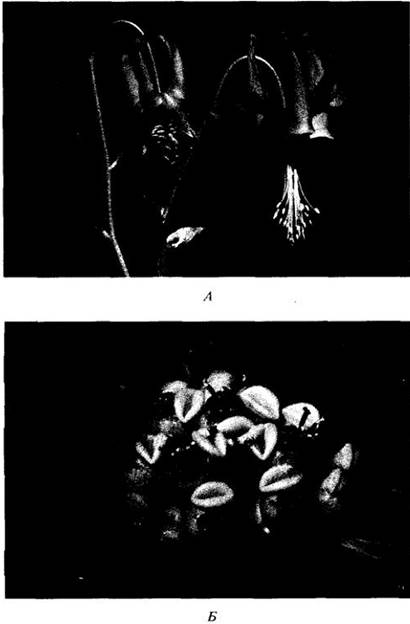
Рис. 29-1. А. Покрытый пыльцой жук- усач (сем. Cerambycidae) на цветках представителя зонтичных (Apiaceae) в горах северо-восточной Аризоны. Б. Надкрылья жука из отложений с Archaeanthus, покрытосеменным, вымершим 95 — 98 млн. лет назад (см. приложение 1 к гл. 29). Эволюция цветковых растений — это в значительной степени история возрастающей специализации взаимоотношений между цветками и насекомыми-опылителями, на первых этапах которой важную роль играли жуки

Чарльз Дарвин в письме к другу назвал внезапное появление представителей этой крупной группы в ископаемых находках «ужасной тайной». В древнейших отложениях, содержащих растительные остатки, встречаются примитивные сосудистые формы с очень простым строением. Затем в девонском и в каменноугольном периодах быстро распространяются папоротники, плауны, хвощи и прогимноспермы. Первые семенные растения появляются в позднем девоне, а в мезозое голосеменные уже преобладают. Наконец, в первой половине мелового периода, т. е. к концу мезозойской эры, возникают покрытосеменные, или цветковые, постепенно захватывающие господствующее положение на всей планете. Около 75 млн. лет назад уже существовали многие современные семейства и даже некоторые ныне живущие роды этого отдела (рис. 29-2).
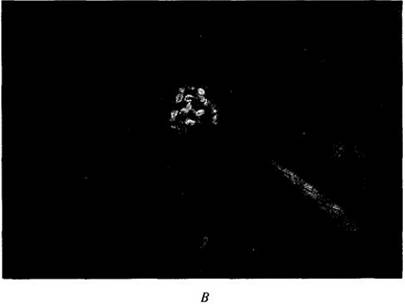
Рис. 29-2. Ископаемые остатки голосеменного (А) и двух покрытосеменных (Б, В) из позднемеловых отложений (около 70 млн. лет назад) Вайоминга. А. Ветвь и отдельные чешуи шишек Araucarites longifolia, вымершего хвойного из семейства араукариевых (A rauca riaceae), распространенного в наше время только в Южном полушарии. К часто разводимым видам этого семейства относится араукария разнолистная (Araucaria heterophylla). Б. Лист вымершей веерной пальмы Sabaliies montana, отдаленно связанной с сабалями юго-востока США. В. Лист вымершего вида Viburnum marginatum из широко распространенного рода кустарников калина

Что же позволило покрытосеменным занять господствующее положение в растительном мире и достичь на сегодняшний день такого поразительного разнообразия? В данной главе мы попытаемся ответить на этот вопрос, сосредоточив внимание на трех моментах: происхождении этой группы, эволюции цветка и роли некоторых химических веществ в эволюции покрытосеменных. Тем самым мы проиллюстрируем процесс эволюции, который начали описывать в предыдущей главе.
Происхождение покрытосеменных
Теперь почти все признают, что цветковые возникли из некоторых примитивных, скорее всего кустарниковых голосеменных.
В меловых остатках вероятных предков покрытосеменных не обнаружено, но ряд более ранних мезозойских и палеозойских голосеменных уже имел определенные сочетания признаков, близких к характерным чертам цветковых. Это наводит на мысль о появлении покрытосеменных раньше того времени, которым датируются их древнейшие ископаемые находки.
Первые остатки, которые с уверенностью можно отнести к покрытосеменным, соответствуют раннему мелу (порядка 125 млн. лет назад) и представляют собой пыльцевые зерна с одной порой. Они похожи на споры папоротников и пыльцу голосеменных, но вполне отличимы от них. По-видимому, пыльцу большинства более ранних покрытосеменных распознать среди остатков других групп уже невозможно. У всех однодольных и большинства примитивных двудольных она похожа на пыльцу первых известных цветковых. В слоях возрастом не менее 120 млн. лет появляются трехпоровые пыльцевые зерна, характерные для всех двудольных, кроме самых примитивных. А уже 80 — 90 млн. лет назад повсюду в мире покрытосеменные были более многочисленны, чем представители любой другой группы растений. Что же мы знаем об их происхождении?
Родственные связи покрытосеменных
Со времен Дарвина ученые пытались определить предков покрытосеменных. Одним из подходов к решению этой задачи был их поиск среди ископаемых остатков семенных растений. При этом особое значение придавалось тому, насколько легко может произойти предполагаемое превращение структур, несущих семязачатки у голосеменных, в плодолистики. В последнее время был предложен иной подход — постараться вместо поиска предков просто выделить главные естественные группы семенных растений и проследить их родство между собой. По степени родства можно затем судить о том, насколько давно они произошли от общего предка. Во главе этого направления сейчас стоит П. Крейн из Музея естественной истории Филда в Чикаго, предоставивший много данных, использованных при составлении нашего раздела.
В простейшем случае для обсуждения филогенетических (эволюционных) связей привлекаются всего три различные группы растений. У папоротников, саговников и сосен, например, ксилема состоит из трахеид, и это наводит на мысль, что когда-то, в далеком прошлом, они имели общего предка (также с трахеидами). Можно поставить вопрос, какие две из этих трех групп разделились позже? В данном случае у нас только три возможности, графически изображаемые тремя схемами (рис. 29-3). Саговники и сосны имеют семена и древесину (вторичную ксилему); ни того ни другого нет у папоротников. Значит, можно предположить, что общий предок саговников и сосен существовал позже, чем общий предок каждой из этих групп и папоротников (рис. 29- 3, А). Признаки, считающиеся унаследованными от общего предка (например, семена, древесина, трахеиды), называются гомологичными. Если ввести в схему дополнительную группу растений, например, дугласию, для выяснения ее родства с саговниками и соснами, потребуется найти более узкие гомологии. Общим для сосен и дугласий является наличие смоляных ходов в листьях; это позволяет предполагать, что оба рода разошлись в эволюции позже, чем каждый из них с саговниками (рис. 29-4). При расширении такого рода анализа за счет увеличения числа признаков и групп растений выводы часто получаются иными, чем при более ограниченном объеме привлекаемых данных. В этих случаях должно быть принято простейшее объяснение связей, т. с. такое, в пользу которого говорит наибольшее число гомологий. Решение может быть проверено, а иногда и изменено при рассмотрении дополнительных признаков и групп растений.
Рис. 29-3. Три варианта возможных родственных связей между папоротниками, саговниками и соснами. Вариант А, вероятно, правильный; он основан на указанных признаках
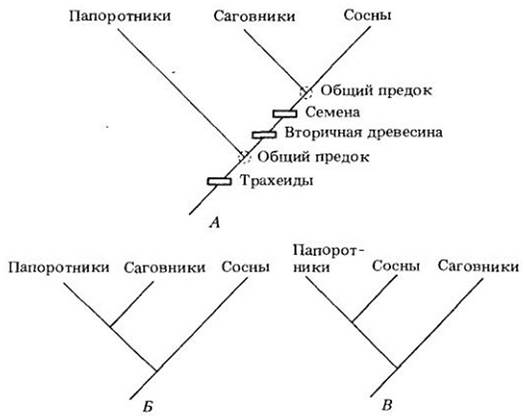
Рис. 29-4. Филогенетические отношения между папоротниками, саговниками, соснами и дугласией с указанием общих признаков, положенных в основу этой конкретной схемы, и примерного времени расхождения групп
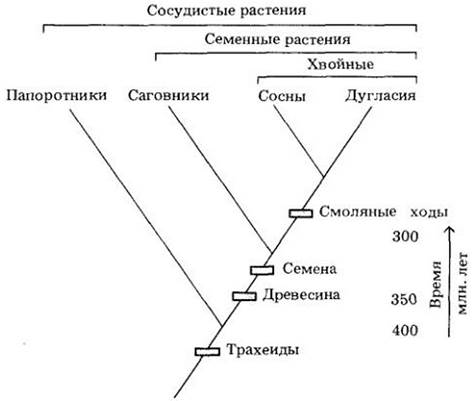
Выяснение таким способом родства приводит к созданию схем, в которых крупные (высшие) группы объединяют несколько более мелких (низших). Это называется иерархией уровней. Каждая группа, или кладон, характеризуется одной или несколькими гомологиями и включает всех потомков одного гипотетического общего предка. Графические изображения родственных связей между такими группами в виде систем ветвей называются кладограммами. Сосудистые растения — это кладон, в пределах которого семенные образуют кладон низшего порядка, а хвойные внутри семенных — кладон еще более низшего (рис. 29-4). Такая схема одновременно предсказывает возможную последовательность появления гомологий в ископаемых остатках. В нашем простом примере палеоботанические данные соответствуют кладограмме: трахеиды впервые появились не позже 400 млн. лет назад, семена и древесина — около 350 млн. лет назад, а смоляные ходы примерно на 50 млн. лет позднее.
В некоторых случаях сходство, кажущееся на первый взгляд признаком родства, не отражает эволюционных связей, изображаемых кладограммой. Например, женские шишки саговников, с одной стороны, и сосен и дугласий с другой — очень похожи, но это сходство лишь внешнее. У саговников каждая их чешуя — просто видоизмененный лист, несущий семя, а у хвойных — сложная структура (см. гл. 18). Таким образом, шишки саговников и хвойных, по- видимому, не произошли от общего предка и не могут быть использованы для характеристики кладона. Такие образования называют аналогичными. Их внешнее сходство обусловлено конвергентной эволюцией (см. с. 72). И напротив, сложные женские шишки сосны и дугласии гомологичны и являются признаком кладона хвойных.
Еще один потенциальный источник путаницы при установлении сходства — слишком узкое понимание частной гомологии. Например, у плаунов, хвощей и папоротников споры свободные (т. е. они рассеиваются, а нс остаются на родительском спорофите), но этот признак не может быть использован для выделения кладона, так как споры этих растений одновременно гомологичны части семени семенных растений. Группа, характеризующаяся свободными спорами, была бы неполной и не включала бы всех потомков одного общего предка, который, по-видимому, дал начало и семенным растениям. В этом примере две гомологии: наличие спор и наличие мегаспор, видоизмененных в семена; они соответствуют двум кладонам, один из которых является частью другого, а сосудистые растения со свободными спорами сами по себе кладона не образуют.
Рассмотренный подход, известный как филогенетический анализ, или кладистика, отодвигает проблему выяснения предков цветковых растений (которые, возможно, еще не обнаружены или вообще не сохранились в ископаемых остатках) на второй план. Его цель — определение главных кладонов семенных растений и оценка их родства с точки зрения относительного времени существования общих предков.
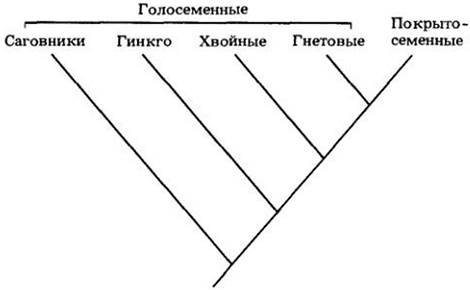
Анализ лучше всего начинать с современных семенных, как правило, лучше изученных, чем родственные им ископаемые. Одна недавно предложенная кладограмма представлена на рис. 29-5. Из нее четко следует, что голосеменные не образуют отдельный кладон и что признаки, которые могли бы характеризовать этот отдел, по-видимому, свойственны и цветковым. Действительно, голосеменные — это просто то, что остается от кладона семенных растений после обособления покрытосеменных.
Рис. 29-5. Филогенетические отношения между четырьмя отделами современных голосеменных и покрытосеменными
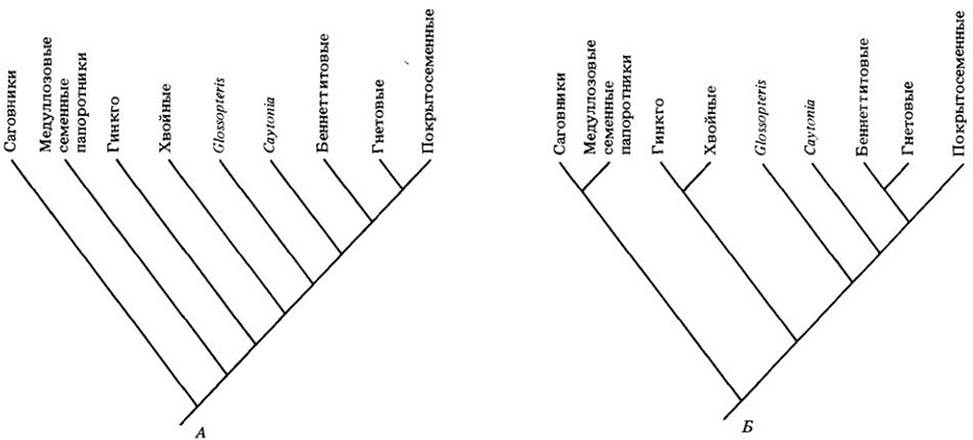
Следующий шаг после анализа современных растений — привлечение данных по ископаемым семенным. Если многие из них удается поместить в группы, известные по современным формам, другие представляют крупные вымершие кладоны. Последние могут играть важную роль при поиске гомологий и тем самым быть чрезвычайно полезными для выяснения эволюционных связей. На рис. 29-6 приводятся две недавно предложенные кладограммы, отражающие родство основных ныне живущих и ископаемых семенных растений. Конечно, эти схемы еще предстоит уточнить в ходе будущих исследований, особенно за счет привлечения новых данных по некоторым важнейшим вымершим группам, но и в таком виде они отражают по нынешним представлениям место цветковых растений в эволюции семенных.
Рис. 29-6. Две возможные схемы филогенетических связей между отделами современных семенных растений и некоторыми родственными группами. Эти кладограммы построены П. Крейном из Музея естественной истории в Чикаго
Эволюционная радиация покрытосеменных
Несколько лет тому назад палеоботаник Д. Акселрод из Калифорнийского университета в Дейвисе высказал мысль, что ранняя эволюция покрытосеменных могла идти далеко от тех равнинных бассейнов, где обычно находят их ископаемые остатки. Вполне вероятно, что на холмах и возвышенностях в тропиках цветковые дали широкий спектр форм, которые имели мало шансов сохраниться в геологических отложениях. По мнению Акселрода, покрытосеменные заняли доминирующее положение в растительности планеты примерно 120 млн. лет назад, когда спустились на равнины.
Древние цветковые обладали многими адаптивными признаками, делающими их особенно устойчивыми к засухе и холоду. Среди них кожистые, обычно мелкие листья, членики сосудов (хорошо проводящие воду клетки) и жесткая, прочная семенная кожура, защищающая молодой зародыш от высыхания. Эти признаки присутствуют не у всех цветковых и отмечаются не только у них, но именно они сыграли главную роль в дифференциации этого отдела.
Важное значение для первоначального и дальнейшего успеха покрытосеменных, по-видимому, имел ряд других факторов. В этой главе мы еще обсудим некоторые из них более подробно. Появление члеников ситовидных трубок повысило эффективность перемещения сахаров по флоэме, а в ксилеме более совершенные проводящие элементы — членики сосудов — заменили трахеиды. Может быть, еще большую роль сыграли характерные для этой группы тонко отлаженные системы опыления и распространения семян, сделавшие возможным существование далеко удаленных друг от друга особей в самых различных местообитаниях. Огромное химическое разнообразие покрытосеменных, содержащих многие вещества, защищающие их от болезней и фитофагов, по-видимому, также имело большое значение (см. ниже). Эти и другие особенности имеют прямое отношение к более быстрому и эффективному размножению покрытосеменных по сравнению с другими растениями.
Важным новоприобретением цветковых была листоиадность. Среди голосеменных также встречаются листопадные формы, например, лиственница и болотный кипарис, но по сравнению с общим объемом группы их крайне мало. Среди покрытосеменных таких растений очень много. Листопадность, по-видимому, впервые возникла в тропических областях с периодической засухой. Потомки первых листопадных растений распространились к северу, где часть года так холодно, что вода для роста недоступна. Другой полезной модификацией цветковых было появление травянистых многолетников, а затем и однолетников, что сделало возможным выживание в более суровых условиях, чем тс, которые переносили их древесные предки. Все эти приспособления оказались особенно существенными при климатических стрессах последних 50 млн. лет истории Земли. Этот период сыграл важнейшую роль в эволюции цветковых.
Около 125 млн. лет назад (возраст первых находок пыльцы покрытосеменных в геологических отложениях) Африка и Южная Америка были соединены между собой, а также с Антарктидой, Индией и Австралией в огромный южный суперконтинент, названный Гондваной (рис. 29-7). Примерно в это время Африка и Южная Америка начали расходиться с образованием южной части Атлантического океана, но в тропической зоне они не полностью разъединились и 90 млн. лет назад. Тогда же Индия начала перемещаться к северу, и около 45 млн. лет назад столкнулась с Азией, что послужило причиной поднятия Гималаев. Приблизительно 55 млн. лет назад от Антарктиды стала отделяться Австралия, но их полное расхождение произошло позже.
Рис. 29-7. Положение материков в то время, к которому относятся первые ископаемые остатки покрытосеменных. В середине мелового периода, около 100 млн. лет назад. Южная Америка была соединена с Африкой, Мадагаскаром и Индией и через Антарктиду с Австралией. Эта объединенная масса суши, выделенная на карте коричневатым цветом, составляла суперконтинент Гондвану
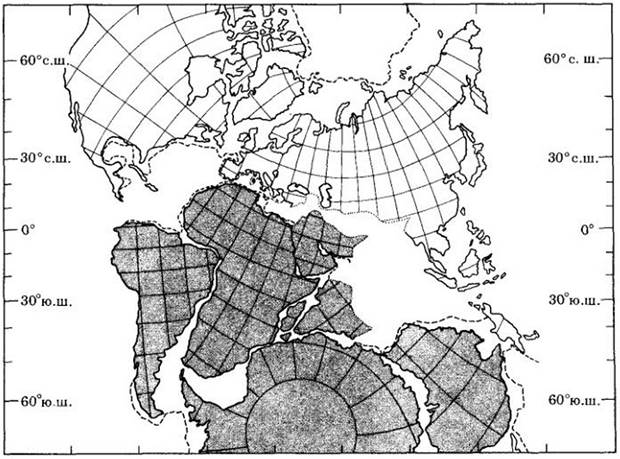
В центральных областях западной Гондваны, соответствующих современным Африке и Южной Америке, по- видимому, были хорошо представлены различные местообитания, от аридного до субгумидного типов, являющиеся, по мнению Акселрода и др., важным центром ранней эволюции покрытосеменных. Ко времени окончательного расхождения двух этих континентов, примерно тогда, когда покрытосеменные, судя по их остаткам, уже в изобилии распространились по всему свету, климат планеты сильно изменился, особенно в экваториальных областях, где он стал мягче, с менее резкими перепадами температуры и влажности. Считают, что с этим связан эволюционный успех древних покрытосеменных, которые усилили свое господство во флорах земного шара.
Когда Индия и Австралия находились на крайнем юге, их растительность была типичной для умеренно-прохладного климата. Ареал некоторых уникальных групп растений и животных, общих в настоящее время для юга Южной Америки и юго-восточной Австралии-Тасмании, сформировался в результате их миграции через Антарктиду задолго до ее полного оледенения, происшедшего около 20 млн. лет назад (рис. 29-8). Когда в последние 55 млн. лет Австралия перемещалась к северу, она попала в широкую зону аридности, граничащую с тропиками; именно тогда на ее территории сильно распространились столь характерные для этого континента пустынные растительные сообщества. Приблизившись к Азии, северная оконечность Австралии оказалась в тропическом климате, и сюда проникли растения и животные тропической Азии. Однако азиатские и австралийские организмы остались в значительной степени географически обособленными; граница, разделяющая области преобладания тех или других, получила название «линия Уоллеса» в честь Альфреда Рассела Уоллеса, одного из первых натуралистов, исследовавших эти территории (он также известен тем, что одновременно с Дарвином предложил теорию эволюции путем естественного отбора). Исходные растения и животные умеренно-прохладной Австралии сохранились на юго-восточной оконечности этого континента, на Тасмании и в Новой Зеландии, отделившейся от Австрало-Антарктиды около 80 млн. лет назад и сместившейся к северо-востоку.
Рис. 29-8. А. Южный бук (Nothofagus menziesii) в верхней долине реки Кэплс (остров Южный, Новая Зеландия) — реликт лесов умеренно-прохладного климата, охватывавших территорию от юга Южной Америки через Антарктиду до Австралии и Новой Зеландии примерно 80 — 30 млн. лет назад. Все это время их ареал дробился на части, разрывы между которыми продолжают расширяться до наших дней. Б. Пальма Nypa fruticans на краю литорального болота. Около 40 — 50 млн. лет назад, когда Африка и Южная Америка тесносоприкасались, этот род был широко распространен по всем теплым областям земного шара, но его ареал постепенно сокращался и теперь ограничен юго-востоком Азии, севером и северо-востоком Австралии, и некоторыми островами в Тихом океане

Первоначальные флора и фауна Индии сохранились далеко не так хорошо, и лишь немногие их реликты дожили до наших дней. Индия «продрейфовала» гораздо дальше, чем Австралия, и пересекла при этом южную аридную зону, тропики, и северную аридную зону. В результате почти все гондванские организмы вымерли; их сменили растения и животные пустынь, тропиков и гор Евразии.
Эволюция цветка
Части цветка
Единственный признак, отличающий покрытосеменные от других групп, — это цветок, структура, довольно подробно обсуждавшаяся в гл. 18. Как уже отмечалось, он представляет собой укороченный побег с различными листоподобными придатками. Какое отношение к листьям имеют части цветка, будет рассмотрено ниже. Большинство наших выводов о его эволюции основано на сравнительном изучении современных форм, хотя значение ископаемых цветков для понимания истории покрытосеменных все возрастает по мерс увеличения числа их находок.
Плодолистик
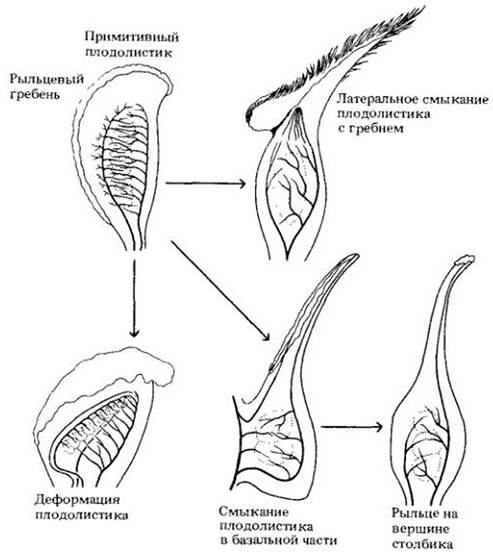
Плодолистик (карпель) в простейшем случае представляет собой сложенную листовую пластинку. Как показано в несколько обобщенном виде на рис. 29-9, специальных мест для улавливания пыльцевых зерен здесь нет. Оба края сложенной пластинки покрыты рыльцевыми волосками. Такой плодолистик охватывает семяпочки, расположенные на его внутренней поверхности. У всех современных покрытосеменных карпели замкнутые, хотя по отдельным группам можно проследить разные стадии этого смыкания. У примитивных цветковых обширные рыльцевые поверхности идут вдоль краев плодолистиков. Более специализированные группы (почти все ныне живущие) имеют рыльца гораздо меньшего размера, расположенные на столбике над завязью.
Рис. 29-9. Некоторые изменения, происходившие в ходе эволюции плодолистика. Примитивный плодолистик сложен вдоль оси. Его пластинка заключает много семязачатков, прикрепленных к ее внутренней поверхности. Края не срослись, а рыльцевая поверхность не имеет четких границ, хотя рыльцевые волоски сильно развиты на гребне, т. е. краях плодолистика. У плодолистиков, происшедших от этого примитивного типа, края в разной степени срастаются, а пыльцеулавливающая рыльцевая поверхность занимает только часть гребня
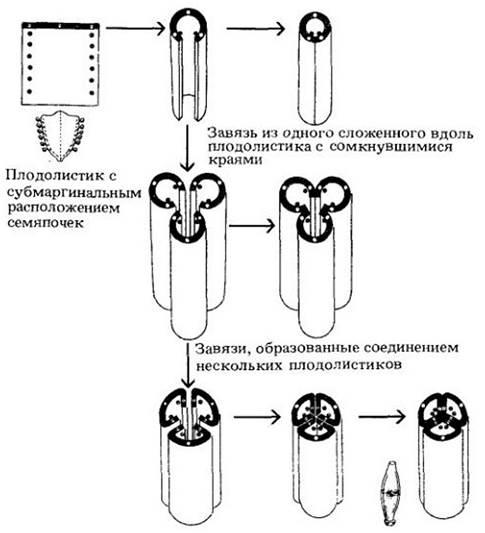
У первых покрытосеменных семяпочки, вероятно, располагались рядами вдоль краев плодолистика на его внутренней поверхности. У более продвинутых групп их размещение стало разнообразнее. У примитивных цветковых было много семяпочек, у более продвинутых их относительно мало. Гинецей в цветке древних покрытосеменных состоял из большого числа отдельных плодолистиков; в ходе эволюционной специализации их число уменьшалось, и они срастались друг с другом (рис. 29-10).
Рис. 29-10. Предполагаемый ход эволюции гинецея
Андроцей
Хотя тычинки современных растений (см., например, рис. 18-39) редко напоминают листья, некоторое сходство этих структур можно обнаружить у магнолий и родственных им растений. Как и у них, примитивные тычинки могли иметь плоскую форму со спорангиями у центра пластинки (рис. 29-11). По одной теории пластинка дифференцировалась в тонкую ножку (тычиночную нить) со спорангиями у верхушки; по другой, не менее приемлемой — тычинки возникли из систем тонких веточек с терминальными спорангиями (веточки постепенно сливались и в некоторых случаях становились листоподобными).
Рис. 29-11. Тычинки примитивных покрытосеменных. У этих древесных растений пыльники образуются на нижней или верхней поверхности листовидных микроспорофиллов. Первый вариант встречается в родах Himantandra и Degeneria (вид снизу), второй — у Austrobaileya и Magnolia (вид сверху). У большинства современных покрытосеменных в тычинках, уже не являющихся листовидными, гораздо меньше стерильной ткани и пыльники находятся на конце тонкой тычиночной нити. Эти различия трудно объяснить, что привело к гипотезе о том, что листовидные тычинки, подобные показанным здесь, могли образоваться путем срастания систем веточек с терминальными спорангиями

У ряда специализированных цветков тычинки, как и плодолистики, срастались. При этом возникали, например, колончатые структуры, как у бобовых, тыквенных, мальвовых (см. рис. 29-13, Г) и сложноцветных. Иногда происходило их прирастание к венчику, в частности в семействах синюховых, норичниковых и губоцветных.
В некоторых эволюционно продвинутых цветках тычинки стали вторично стерильными, т. е. утратили спорангии и превратились в специализированные структуры, например, в нектарники — железы, выделяющие нектар, сахаристую жидкость, привлекающую опылителей и служащую им пищей. (Важно отметить, однако, что в большинстве случаев нектарники не являются видоизмененными тычинками, а образуются за счет других частей цветка.) Ниже будет показано, что тычинки играли определенную роль и в эволюции лепестков.
Околоцветник
Околоцветник состоит из чашелистиков и лепестков. Первые у большинства растений зеленые и фотосинтезирующие. Этим, а также тем, что в них заходит столько же проводящих пучков, сколько и в листья данного вида (часто, хотя и нс всегда, более одного), чашелистики подобны листьям, и считается. что они произошли непосредственно от них.
В немногих семействах, например, у кувшинковых, лепестки, по-видимому, возникли из чашелистиков. Однако у большинства покрытосеменных в лепестки, вероятно, видоизменились тычинки, утратившие перед этим спорангии и приобретшие в результате новую функцию — привлечения к цветку потенциальных опылителей. Большинство лепестков, как и тычинки, снабжается одним проводящим пучком, отличаясь, таким образом, от чашелистиков, в которых, как уже упоминалось, обычно столько же пучков, что и в листьях того же вида (часто три или более). И в чашелистиках, и в лепестках проводящие пучки обычно разветвляются, поэтому сколько их заходит в орган, определить по числу жилок на пластинке нельзя.
В ходе эволюции многих групп покрытосеменных происходило срастание лепестков с образованием характерного для многих семейств трубчатого венчика. При этом с ним часто срастались тычинки (как бы вырастающие из венчика). В ряде эволюционно продвинутых семейств чашелистики также слились в трубку.
Направления эволюции цветка
Среди современных растений цветком, наиболее близким по общему строению к примитивному, обладают магнолиевые (рис. 29-12). У них он включает многочисленные плодолистики, тычинки и части околоцветника, причем все эти структуры четко обособлены друг от друга. Кроме того, еще хорошо заметно их спиральное расположение на коническом цветоложе. Однако сильная его вытянутость и «шишковидная» упаковка плодолистиков явно вторичны, отражают специализацию магнолиевых и не характерны для других примитивных покрытосеменных.
Рис. 29-12. Строение цветка магнолии крупноцветковой (Magnolia grandiflora). На коническом цветоложе много спирально расположенных плодолистиков с отогнутыми столбиками. Под ними — светло-кремовые тычинки (А, Б). Нераскрывшийся бутон; спереди части околоцветника удалены, чтобы показать рыльца и тычинки, еще не рассеивающие пыльцу. Б. Ось цветка на второй день цветения; рыльца уже невосприимчивы, а тычинки пылят. В. Плоды; видны плодолистики и ярко- красные семена на тонких ножках

Сравнивая такие цветки с более специализированными, можно проследить четыре главных направления эволюции этой структуры (рис. 29-13, см. также рис. 18-43).
1. От большого и нефиксированного числа частей к малому и четко определенному.
2. От четырех типов частей до трех, двух, а иногда и до одного у более продвинутых групп. Побег укорачивается, и их исходное спиральное расположение перестает замечаться. Части цветка срастаются.
3. От верхней завязи к нижней.
4. От радиальной симметрии, или актиноморфности, к билатеральной, или зигоморфности.
Рис. 29-13. Примеры специализированных цветков. А. Зимолюбка зонтичная (Chimaphila umbellata); число чашелистиков (не видны) и лепестков уменьшилось до 5, тычинок — до 10, а 5 плодолистиков объединившись в один сложный гинецей с одним рыльцем. Б. Лотос желтый (Nelumbo lutea). Недифференцированные чашелистики и многочисленные лепестки, а также тычинки расположены по спирали; плодолистики слились в сложный гинецей. В. Жимолость Lonicera hispidula. Завязь нижняя, двух- или трехгнездная; чашелистики редуцированы до мелких зубчиков на ее апексе. Лепестки объединены в трубку венчика зигоморфного (билатерально-симметричного) цветка. Высовывающиеся из трубки 5 тычинок прикреплены к внутренней стенке трубки. Столбик длиннее тычинок, и рыльце возвышается над ними. Прежде чем опылитель, посещающий этот цветок, достигнет его пыльников, он должен коснуться рыльца, оставив на нем принесенную с другого цветка пыльцу. Плоды этого растения показаны на рис. 29-46. В. Г. Схема цветка хлопчатника (Gossipium) с тычинками, объединенными вокруг столбика в колонку


Примеры специализированных семейств
К группам с наиболее специализировавшимися в ходе эволюции цветками принадлежат сложноцветные (Astcraceae, или Compositac) среди двудольных и охридные (Orchidaceae) из однодольных.
Сложноцветные
У растений этого семейства эпигинные цветки сравнительно мелкие и тесно скученные в соцветие корзинку. Их нижняя завязь одногнездная и образована двумя сросшимися плодолистиками с единственным семязачатком (рис. 29-14).
Рис. 29-14. Сложноцветные (Asteraceae). А. Схема корзинки представителя этого семейства. Устройство отдельных цветков подчинено общему плану строения корзинки, функционирующей как один крупный, привлекающий опылителей цветок. Б. Бодяк Cirsium pastoris. Виды его трибы имеют только трубчатые цветки. У этого бодяка они ярко-красные и регулярно посещаются колибри, главными и агентами его опыления. В. Agoseris, родственный одуванчику (Taraxacum). В соцветиях представителей этой трибы (группы сложноцветных, к которым относятся одуванчик, цикорий и родственные им виды) трубчатых цветков нет, развиты только язычковые, более крупные по краю соцветия. Г. Подсолнечник (Helianthus annuus)

Число тычинок в цветке сведено до пяти, они обычно сращены друг с другом и с венчиком. Лепестков тоже пять, сросшихся друг с другом и с завязью, а чашелистики отсутствуют или редуцированы до рядов щетинок или чешуи так называемого паппуса. Он часто способствует распространению семян ветром, например, у относящегося к сложноцветным, одуванчика (рис. 29-14, В; см. также рис. 29-42). У других представителей этого семейства, например, у череды (Bidens), паппус может нести крючки, служащие для прикрепления плода к покровам животных, что повышает вероятность его переноса на новое место. У многих сложноцветных в корзинке два типа цветков: трубчатые в центральной части и язычковые по краю. Последние часто имеют завязь, но иногда полностью стерильны. У некоторых сложноцветных, например, маргаритки, подсолнечника или ромашки, их сросшийся венчик имеет вид длинного «лепестка».
Вообще корзинка сложноцветных напоминает крупный одиночный цветок, однако в отличие от него созревает в течение нескольких дней — отдельные цветки раскрываются в центростремительной последовательности. Вследствие этого семязачатки одной корзинки могут быть оплодотворены пыльцой из разных источников. Успех такой эволюционной стратегии доказан многочисленностью этого, вероятно, самого крупного семейства цветковых, в котором около 22 тыс. видов.
Орхидные
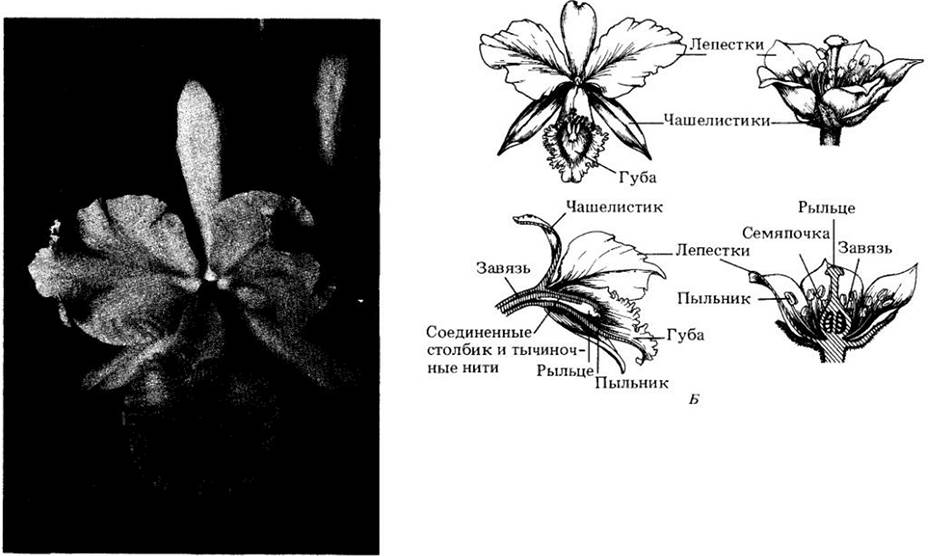
Другой, очень удачный, план строения цветка демонстрируют орхидные, в противоположность сложноцветным относящиеся к однодольным. Их не менее 17 тыс. видов, но классификация многих родов слабо разработана. Во всяком случае, это также одно из крупнейших семейств. Большинство его видов тропические; только у примерно 140 ареал захватывает США и Канаду. Три плодолистика орхидных, как и у сложноцветных, сросшиеся, завязь нижняя (рис. 29-15), однако в завязи многие тысячи крошечных семяпочек, и соответственно опыление может приводить к образованию огромного числа семян. Обычно присутствует только одна тычинка (в подсемействе Diandra — две), соединенная со столбиком и рыльцем в сложную структуру — колонку. Все содержимое пыльника удерживается вместе и переносится как единое целое — поллиний (см. рис. 29-26, Б). Из трех лепестков два боковых видоизменены в крылья, а третий в чашеобразную губу, часто очень крупную и яркую. Чашелистиков также три, часто окрашенных и похожих на лепестки. Цветки всегда билатерально-симметричны и во многих случаях имеют причудливую внешность.
Рис. 29-15. А. Орхидея 143 рода Саttlеуа. Орхидные — одно из наиболее специализированных семейств однодольных. Б. Сравнение частей цветка орхидеи (слева) и радиально-симметричного цветка (справа). «Губа» — видоизмененный лепесток, служащий посадочной площадкой для насекомых
У орхидных встречаются цветки величиной с булавочную головку и диаметром более 20 см. В некоторых родах известны сапрофитные растения; два австралийских вида ведут полностью подземный образ жизни, и их цветки появляются в трещинах почвы, где опыляются мухами (рис. 29-16). При коммерческом разведении орхидей их клонируют, разделяя меристематическую ткань и быстро получая таким образом тысячи одинаковых растений. В этом семействе зарегистрировано более 60 тыс. гибридов, причем многие из них между двумя или более родами. Плоды рода Vanilla — естественный источник популярной пряности ванили (рис. 29-17).
Рис. 29-16. Rhizanthella — орхидея из Западной Австралии, растущая целиком под землей. В трещинах почвы, образующихся во время сухого сезона, можно видеть ее цветки, никогда не выходящие на поверхность. А. Вид сверху (мусор и листья расчищены): три расходящиеся брактеи, между которыми пролезают опылители (мухи). Б. Цветки, окруженные защищающими их брактеями

Рис. 29-17. Ваниль (Vanilla) —орхидея, из которой в промышленных масштабах получают одноименную пряность. Первыми ее начали использовать ацтеки на территории теперешней Мексики. Теперь ваниль разводят в основном на Мадагаскаре, других островах западной части Индийского океана и вообще в Старом Свете. Шоколад — это смесь какао с ванилью. Синтетическое ароматическое начало ванили — ванилин — широко используется сейчас вместо натурального продукта, экстрагируемого из высушенных и сброженных плодов этой орхидеи. А. Цветки ванили (Vanilla planifolia). Б. Искусственное опыление ванили в Мексике; оно применяется даже у диких растений, чтобы обеспечить хороший урожай плодов, из которых экстрагируется пряность
Агенты эволюции
В отличие от животных растения нс могут перемещаться с места на место в поисках пищи, убежища или партнера для спаривания и вынуждены удовлетворять эти свои «потребности» с помощью ростовых процессов и образуемых структур. Однако у покрытосеменных сформировался набор признаков, обеспечивающих определенную активность в поисках полового партнера; это — особенности цветка. Привлекая им насекомых и других животных и направляя их действия так, чтобы повысить вероятность перекрестного опыления (т. е. спаривания неблизкородственных организмов), покрытосеменные в определенном смысле являются не менее подвижными, чем животные. Как это было достигнуто?
Первые семенные растения — различные группы голосеменных — опылялись пассивно. Их пыльца разносилась ветром и лишь случайно оказывалась около семязачатков. Находясь на листьях или внутри шишек, тс выделяли из своего микропиле капли клейкого сока. Они служили для улавливания пыльцевых зерен и втягивания их в микропиле, как и у современных голосеменных. Насекомым (вероятно, это были жуки, см. рис. 29-1), питавшимся соком и смолой на стеблях и листьях, попадались богатые белками пыльцевые зерна и клейкие капли, выделенные семязачатками. Регулярно используя эти новые источники пищи, насекомые в результате своих посещений стали переносить пыльцу с одного растения на другое. Такой способ опыления, очевидно, сразу же оказался более эффективным, чем пассивный, т. е. с помощью ветра.
Чем привлекательнее были для жуков растения, тем чаще они посещались и тем больше могли образовать семян. Любое изменение фенотипа, способствовавшее повышению частоты или эффективности таких посещений, немедленно создавало селективное преимущество. Это обусловило несколько важных эволюционных изменений. Например, селективное преимущество получили растения с цветками, представлявшими особо ценный источник пищи для опылителей. В дополнение к пыльце, клейкой жидкости вокруг семязачатков и съедобным частям самого цветка в нем образовывались нектарники, секретирующие нектар — питательную сахаристую жидкость, служащую источником энергии для насекомых и других животных.
Привлечение к цветкам опылителей создало новую проблему: необходимость защиты семязачатка от поедания. Возникновение замкнутого плодолистика, вероятно, было одним из путей ее решения. Дальнейшие изменения в строении цветка, например, образование нижней завязи, также могли явиться одним из средств защиты семязачатка.
Другим важным шагом было появление обоеполого цветка. Присутствие в нем и плодолистиков, и тычинок (в противоположность, например, мужским и женским шишкам у хвойных) делало более эффективным каждое посещение опылителя, который теперь одновременно мог и забирать пыльцу данного растения, и оставлять на нем чужую.
В начале третичного периода, т. е. 40 — 60 млн. лет назад, повысилось обилие и разнообразие специализированных групп посещающих цветки насекомых — пчел и бабочек, эволюционировавших к этому времени вместе с покрытосеменными уже в течение почти 50 млн. лет. Рост численности и дифференциация этих животных непосредственно связаны с этими же процессами у покрытосеменных. В свою очередь насекомые оказывали глубокое влияние на эволюцию цветковых, во многом способствуя увеличению их разнообразия.
Если данный вид опыляется насекомыми только одного или немногих видов, ему выгодно специализироваться в соответствии с признаками этих посетителей. Многие изменения примитивного цветка покрытосеменных были специальными адаптациями, способствовавшими «закреплению» определенных опылителей за цветками с особыми признаками.
Ниже будут описаны некоторые эволюционные изменения цветка, возникшие в ходе его приспособления к конкретным опылителям.
Цветки, опыляемые жуками
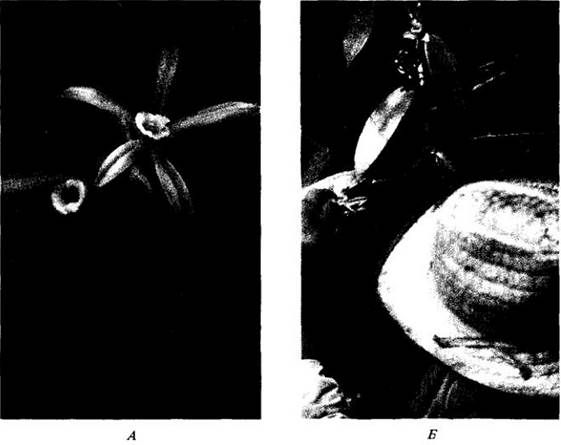
Ряд современных видов покрытосеменных опыляется исключительно или преимущественно жуками (рис. 29-18; см. также рис. 29-1). Цветки у них либо крупные одиночные (как у магнолиевых, некоторых лилейных, калифорнийской эппольции и шиповников), либо мелкие и собранные в соцветие, как у кизила, бузины, спиреи и многих зонтичных (Арiасеае) (см. рис. 29-1, А). Часто посещают цветки представители 16 семейств жуков, хотя, как правило, основной пищей им служат сок вегетативных частей растений, плоды, помет и гниющие остатки. У жуков обоняние развито гораздо сильнее зрения, поэтому опыляемые ими цветки часто белые или неяркие, но имеют сильный запах (рис. 29-19, А), обычно фруктовый, пряный или напоминающий неприятный запах брожения, чем он отличается от сладковатого аромата цветков, опыляемых пчелами и бабочками. Некоторые опыляемые жуками цветки выделяют нектар, в других эти насекомые кормятся непосредственно лепестками или особыми питательными тельцами (скопления клеток, на поверхности различных частей цветка), а также пыльцой. В большинстве случаев семяпочки здесь хорошо защищены завязью и находятся вне досягаемости жующих челюстей опылителей.
Рис. 29-18. Питающийся пыльцой жук (Asclera ruficollis) на открытом блюдцевидном цветке печеночницы американской (Hepática americana). Все виды этого семейства жуков (узкокрылки — Oedemeridae) во взрослом состоянии являются облигатно пыльцеядными

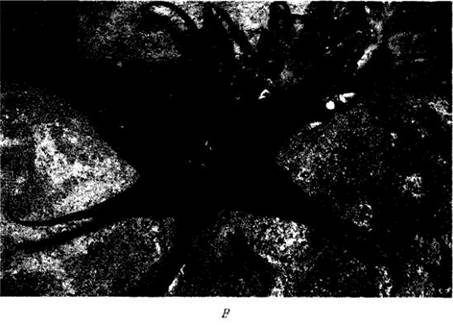
Рис. 29-19. А. «Скунсова капуста» (Lysichiton americanum) опыляется мелкими, активно летающими жучками из семейства стафилинид (Staphylinidae), привлекаемыми очень сильным запахом этого растения. Другие виды того же семейства (Аrасеае) имеют соцветия с запахом, напоминающим запах протухшей рыбы или падали, и опыляются падальными мухами. Б. Одно из таких растений — Symplocarpus foetidus. Ряд видов других семейств имеет близкий запах и также опыляется падальными мухами; хорошими примерами служат Stapeliaschinzii (В) и родственные ей, преимущественно африканские, представители семейства ластовневых

Цветки, опыляемые пчелами, осами и мухами
Пчелы — важнейшая группа животных, посещающих цветки. Они опыляют больше видов растений, чем кто-либо еще. Пчелы живут за счет нектара, а рабочие особи, кроме того, собирают пыльцу на корм личинкам. Их ротовой аппарат, щетинки на теле и другие придатки являются специальными приспособлениями, облегчающими сбор и перенос этих продуктов (рис. 29-20). Как показали Карл фон Фриш и другие исследователи поведения насекомых, пчела быстро научается различать цвета, запахи и внешние очертания предметов, хотя солнечный спектр она, как и большинство насекомых, видит несколько иначе, чем мы. В отличие от человека пчелы воспринимают его ультрафиолетовую часть и не улавливают красную, которая как бы сливается с фоном.
Рис. 29-20. Пчелы так же высокоспециа- лизированы, как и цветки, вместе с которыми они коэволюционировали. Их ротовые придатки преобразовались в сосущую трубку с язычком. Первый членик каждой из трех пар ног несет на своей внутренней поверхности пучок щетинок. На первой и второй парах эти щеточки собирают пыльцу, пристающую к мохнатому телу пчелы. Щеточки третьей пары ног счищают пыльцу со щеточек первых пар и с брюшка. С них пыльца заталкивается в корзиночки, окаймленные волосками выемки на верхнем членике третьей пары ног. На рисунке показана медоносная пчела (Apis mellifera), питающаяся на цветке розмарина лекарственного (Rosmar tus officinalis). Его тычинки и рыльце высовываются из венчика и загибаются вверх, так что соприкасаются с мохнатой спинкой, любой посетившей цветок пчелы подходящего размера; видно, что пыльники оставили на насекомом белые пыльцевые зерна

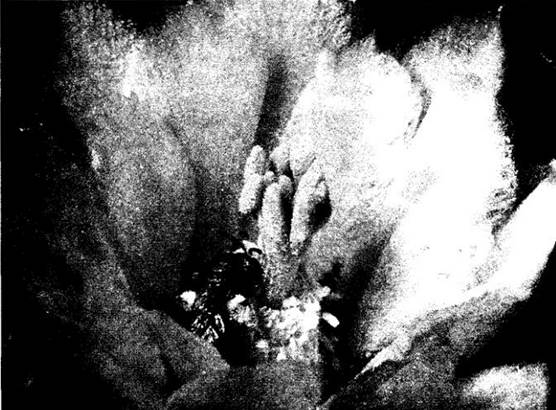
Многие их виды — особенно одиночные, составляющие большую часть этой группы (рис. 29-21), — высоко специализированы в выборе посещаемых цветков. Такое постоянство повышает эффективность отдельного насекомого или данного вида как опылителя. В связи с этой специализацией пчелы с четко фиксированным кормовым поведением часто имеют обусловленные им заметные морфологические и физиологические адаптации. Так, если они посещают растения с крупной пыльцой, то аппарат для ее сбора снабжен грубыми щетинками, а если собирают нектар из цветков с длинной трубкой, то их ротовые придатки удлинены. Достигая такой степени постоянства в своих «привязанностях», пчелы становятся мощным эволюционным фактором, направляющим специализацию посещаемых ими растений. Известно около 20 тыс. видов пчел. Большинство их кормятся на цветках.
Рис. 29-21. Пчела галикт (семейство Halictidae), собирающая пыльцу с тычинок цветка кактуса из рода Еchinocereus (Нижняя Калифорния, Мексика). В центре цветка над тычинками — рыльца
Цветки, коэволюционирующие вместе с пчелами, имеют броские, ярко окрашенные лепестки, обычно голубые или желтые, часто с особым узором, по которому насекомые могут без труда их распознавать. Иногда он включает «нектарный указатель», т. е. специальные знаки, обозначающие положение нектара (рис. 29-22). Эти цветки никогда не бывают чисто красными. Специальные методы фотографирования показали, что часто на них присутствуют характерные метки, невидимые для человека (рис. 29-23).
Рис. 29-22. «Нектарные указатели» на цветках наперстянки (Digitalis purpurea) служат для привлечения насекомых- опылителей. Нижняя губа сростнолепестного венчика образует «посадочную площадку», обычно присутствующую у «пчелиных» цветков

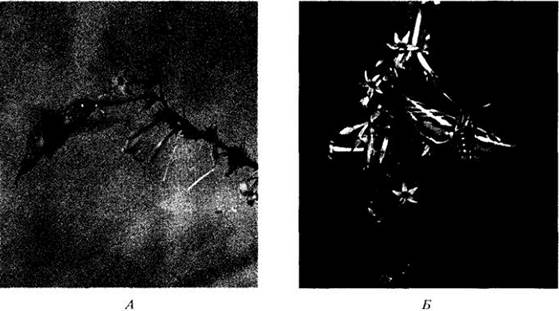
Рис. 29-23. Цветовое восприятие большинства насекомых несколько отличается от человеческого. Например, пчелы четко различают невидимый для человека ультрафиолет. На фотографии показано, что цветок калужницы болотной (Caltha palustris), кажущийся нам ярко-желтым (А), отражает ультрафиолетовые лучи только той своей частью, которая выглядит на рис. Б светлой. Она отражает также желтые лучи, которые, сочетаясь с ультрафиолетовыми, дают так называемый «пчелиный пурпурный» цвет. Темные части цветка поглощают ультрафиолет, поэтому выглядят для пчелы желтыми (см. также с. 217)

Для «пчелиных» цветков типичны нектарники у основания трубки венчика; они часто погружены так, что доступны только для специализированных ротовых придатков пчел и недосягаемы, например, для грызущего аппарата жуков. Такие цветки, как правило, имеют своего рода «посадочные площадки» (см. рис. 29-22).
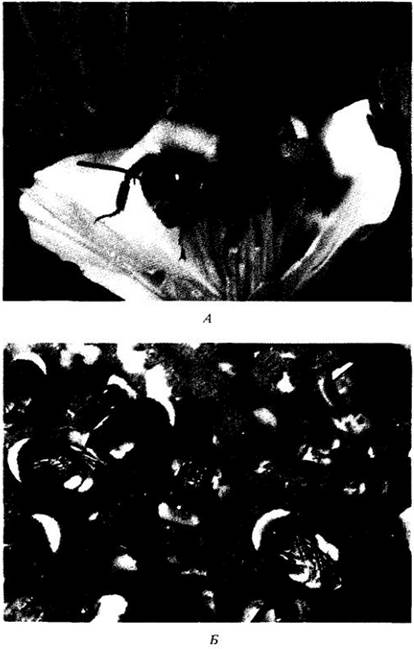
Одни из наиболее обычных посетителей цветков в северном умеренном поясе — шмели (рис. 29-24). Это также общественные перепончатокрылые. Их самки перезимовывают, а весной откладывают яйца, основывая новую колонию. Шмели не способны к полету, пока температура мышц, приводящих к движению их крыльев, не достигнет 32°С; для ее поддержания они должны непрерывно кормиться на богатых нектаром цветках. В прохладных областях Северной Америки и Евразии многие растения, включая люпины, живокость и иван-чай, по всему своему ареалу регулярно опыляются шмелями.
Рис. 29-24. Шмели (Bombus). Эти общественные перепончатокрылые являются важными опылителями многих родов растений в прохладных областях Северного полушария и интродуцированы за пределы своего естественного ареала для опыления таких кормовых растений, как, например, клевер ползучий (Trifolium repens). А. Шмель, собирающий пыльцу на эшшольции калифорнийской (Eschscholzia californica). Б. Часть подземного шмелиного гнезда; видны ячейки, в которых червеобразные личинки завершают свое развитие. Шмели снабжают эти ячейки пыльцой и отрыгиваемым нектаром. Если вся колония шмелей может «обслуживать» в течение сезона цветки самых разных видов, отдельные насекомые нередко посещают цветки только определенных растений, встречающихся по единственному, ведущему от гнезда маршруту
Некоторые наиболее эволюционно продвинутые цветки, особенно у орхидных, образовали сложные проходы и ловушки, заставляющие пчел проникать внутрь них и выходить наружу по определенному пути. В результате пыльник и рыльце прикасаются к телу насекомого в определенной точке и в необходимой последовательности (см. рис. 29-20).
Еще более удивительная стратегия опыления выработалась у орхидей рода офрис (Ophrys). Их цветки очень похожи на самку пчел, ос или мух (рис. 29-25). Самцы этих насекомых начинают летать ранней весной, раньше самок. Одновременно зацветают офрисы, и самцы насекомых пытаются спариваться с их цветками. При этом пол линий может прикрепиться к телу насекомого и, когда оно посетит другой цветок, попасть на рыльце последнего, за счет чего произойдет опыление.
Рис. 29-25. Похожий на пчелу цветок офриса (Ophrys speculum) с Сардинии привлекает самцов пчел, которые настолько обманываются его сходством с самкой своего вида, что пытаются спариться с цветком. При этом к насекомым часто прикрепляются пол линии, переносимые в результате на другие экземпляры офриса

Целый ряд по-разному устроенных цветков опыляется различными двукрылыми, включая комаров. Эти насекомые кормятся нектаром, но не собирают пыльцу и не запасают пищу для своих личинок. Примеры цветков, опыляемых комарами и мухами, приведены на рис. 29-19, Б, В.
Цветки, опыляемые бабочками
Цветки, коэволюционирующие вместе с дневными бабочками, во многих отношениях похожи на цветки, опыляемые пчелами, главным образом тем, что привлекают насекомых также рисунком и запахом. Однако некоторые чешуекрылые способны различать красный цвет, и опыляемые ими цветки бывают красными или оранжевыми.
У видов, как правило опыляемых ночными бабочками, белые или бледноокрашенные очень ароматные цветки, как. например, у некоторых видов табака (Nicotiana), причем их сильный сладкий запах часто проявляется только после захода солнца. Среди таких растений и виды ослинника (Oenothera) с желтыми цветками (см. рис. 9-13, Б), и ярко-розовый амариллис белладонна (Amaryllis belladonna).
В цветках, опыляемых бабочками, нектарники часто находятся в основании длинной узкой трубки венчика или шпорца, откуда его, как правило, могут доставать только эти насекомые с их вытянутым сосущим ротовым аппаратом. Бражники, например, обычно не залезают в цветок, как пчелы, а зависают над ним, вставляя свой длинный хоботок в цветочную трубку. Соответственно эти цветки не имеют «посадочных площадок», ловушек и сложного внутреннего устройства, как иногда наблюдается при опылении пчелами. Среди менее специализированных бабочек-опылителей обычны более мелкие виды, не нуждающиеся в таком количестве энергии, как бражники; посещаемые ими цветки, как правило, мельче, с относительно короткими трубками. Насекомые по ним просто ползают. Один из примеров наиболее тесных взаимоотношений между растением и чешуекрылым показан на рис. 29-28.
Цветки, опыляемые птицами
Некоторые пернатые регулярно посещают цветки, питаясь их частями, нектаром, и живущими здесь насекомыми; многие из этих птиц являются опылителями. В Северной и Южной Америке это в основном колибри, а в других частях земного шара представители других специализированных семейств.
Цветки, опыляемые птицами, выделяют обильный жидкий нектар (у некоторых видов он ко времени созревания пыльцы даже вытекает наружу), однако запах у них слабый, что связано с плохим развитием у пернатых обоняния. Зато они очень хорошо воспринимают цвета (примерно в том же спектральном диапазоне, что и человек), поэтому нс удивительно, что окраска большинства опыляемых ими цветков броская, обычно желтая и красная (см. рис. 29-14, Б), как, например, у водосбора канадского (рис. 29-31, А), фуксии, страстоцвета, эвкалипта, гибискуса, пуансеттии (рис. 28-31, Б, В), многих кактусов, бромелиевых и орхидных. Обычно такие цветки крупные или собраны в мощные соцветия, что связано с необходимостью привлекать пернатых своим видом и вмещать большие количества нектара.
Рис. 29-31. Примеры цветков, опыляемых птицами. А. Водосбор Aquilegia canadensis. С обычными лепестками чередуются элементы околоцветника, видоизмененные в заполненные нектаром трубки. Колибри, посещающие эти поникающие цветки, собирают нектар на лету; для большинства других животных он недосягаем. Б, В — молочай пуансеттия (Euphorbia pulcherrima). У этого известного растения родом из Мексики цветки мелкие зеленоватые и скученные, но каждый из них имеет крупный желтый нектарник, выделяющий обильный нектар. Видоизмененные верхние листья своим ярко-красным цветом привлекают к скученным цветкам колибри
Птицы и другие животные-опылители обычно посещают цветки определенного вида растений в течение коротких промежутков времени, но это только один фактор, способ ствующий ауткроссингу (перекрестному опылению между особями одного вида). Необходимо также, чтобы опылитель не ограничивал свои посещения одним цветком или цветками одного и того же растения. Если на них регулярно кормятся крупные, интенсивно расходующие энергию птицы, бражники, летучие мыши, нектар должен производиться в количествах, достаточных для того, чтобы обеспечить метаболические потребности этих животных и тем самым их неоднократное посещение. Однако, если обильный нектар будет доступен животным, тратящим меньше энергии, например, мелким пчелам или жукам, они, по-видимому, будут задерживаться на одном цветке и, насытившись там, нс станут перелетать на другие растения, осуществляя ауткроссинг. В связи с этим у цветков, регулярно опыляемых крупными видами, в частности колибри, прослеживается тенденция к образованию нектара глубоко в трубках венчика или в других местах, недоступных для более мелких животных. Аналогичным образом красный цвет привлекает птиц, но не различается большинством насекомых. Пернатые в свою очередь, как и люди, слабо воспринимают запахи. Таким образом, непахучие красные цветки плохо заметны насекомым и, как правило, не привлекают их, что является выгодной адаптацией растений, выделяющих большие количества нектара.
Цветки, опыляемые рукокрылыми
Летучие мыши, посещающие цветки, известны в тропических областях Старого и Нового Света. Более 250 видов этих животных (около четверти их общего видового состава) хотя бы в небольшом количестве включают в своей рацион нектар, плоды или пыльцу. Рукокрылые, питающиеся исключительно или в основном на цветках, имеют узкую вытянутую мордочку и длинный, далеко высовывающийся язык, кончик которого иногда напоминает кисточку, а их передние зубы часто редуцируются или утрачиваются совсем.
Цветки, опыляемые летучими мышами и птицами, во многом сходны; они крупные, прочные и производят много нектара (рис. 29-32). Поскольку рукокрылые питаются только ночью, «их» цветки обычно неяркие и часто раскрываются только после захода солнца. Многие из них трубчатые или имеют другие структурные особенности для предохранения нектара. У многих растений, привлекающих летучих мышей для опыления или распространения семян, цветки и плоды либо свисают на длинных цветоножках ниже листвы, где рукокрылым легче летать, либо образуются на стволах. Летучие мыши отыскивают цветки, пользуясь главным образом обонянием, поэтому для последних характерен очень сильный запах брожения или фруктов. Эти животные, перелетая от дерева к дереву, слизывают нектар, поедают части цветка и пыльцу, одновременно перенося ее на своей шерсти с одного растения на другое. Они опыляют и (или) распространяют семена по крайней мере 130 родов покрытосеменных.
Рис. 29-32. Просовывая мордочку в трубчатый венчик цветка кактуса Lemaireocereus, эта летучая мышь из рода Leptonycteris слизывает нектар своим длинным, покрытым щетинками языком. Пыльцевые зерна, осыпавшие голову и шею рукокрылого, переносятся на следующий посещаемый им цветок. Опыляемые летучими мышами цветки имеют тусклый цвет, затхлый запах (похожий на испускаемый этими животными для привлечения друг друга) и раскрываются ночью

Недавно обнаружено, что некоторые летучие мыши получают значительную часть белка из поедаемой пыльцы. Оказалось, что пыльца опыляемых ими растений содержит его гораздо больше, чем у насекомоопыляемых, что служит еще одним примером коэволюции.
Ветроопыляемые растения
На рубеже нашего столетия многие ботаники считали, что цветок ветроопыляемых растений самый примитивный, а все прочие типы произошли от него. У хвойных, которые в то время считались некоторыми учеными прямыми предками покрытосеменных, мелкие невзрачные однополые шишки без запаха опыляются ветром. Многие ветроопыляемые цветки также неяркие, относительно непахучие и не образуют нектара. Лепестки их мелкие или вообще отсутствуют. Такие цветки часто раздельнополы. Однако изучение других признаков ветроопыляемых покрытосеменных (в частности, их специализированной древесины) убедило большинство ботаников в том, что они произошли не от хвойных, а от насекомоопыляемых цветковых.
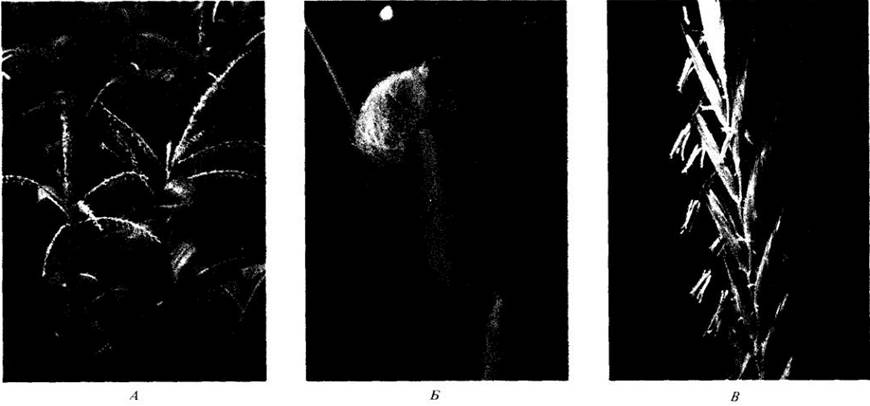
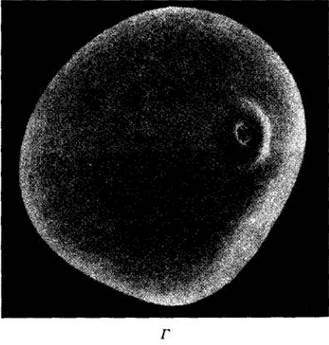
Рис. 29-33. В противоположность большинству покрытосеменных злаки опыляются ветром. У кукурузы (Zea mays) мужские соцветия (метелки) находятся наверху стебля (A), а женские соцветия (Б) с длинными высовывающимися рыльцами — в нижней его части. В. Для злаков характерны крупные перистые рыльца, хорошо улавливающие переносимую ветром пыльцу, рассеиваемую свисающими пыльниками, как показано здесь на примере рода пырей (Agropyron). Г. Фотография пыльцевого зерна кукурузы, сделанная с помощью сканирующего электронного микроскопа; видна его гладкая стенка, свойственная большинству ветроопыляемых растений, и единственная апертура, что типично для однодольных
Согласно современным представлениям, их эволюция шла независимо от нескольких предковых групп. Лучше всего ветроопыляемые виды представлены в умеренных областях и относительно редки в тропиках. В первом случае деревья одного вида часто растут густыми массивами и цветут ранней весной в безлистном состоянии, что облегчает перенос пыльцы ветром. В тропиках на такой же площади встречается гораздо больше пород и экземпляры одного вида могут быть сильно удалены друг от друга. Более того, во многих тропических сообществах деревья вечнозеленые, и поэтому распространение пыльцы ветром не может происходить так же легко, как в листопадных лесах умеренного климата. В этих условиях гораздо эффективнее опыление насекомыми, способными отыскивать растения того же вида,
разделенные в некоторых случаях расстояниями в 20 км и более.
Поскольку ветроопыляемые растения не нуждаются в насекомых для переноса пыльцы, они не расходуют энергию на производство для них пищевого «вознаграждения». Однако ветроопыление очень неэффективно и оправдывает себя только там, где большое число одновидовых особей растет достаточно близко друг к другу. Почти вся подхватываемая ветром пыльца падает на землю в радиусе нескольких сот метров от родительского растения, поэтому, если отдельные экземпляры данного вида удалены друг от друга, шансы достижения пыльцевым зерном рыльца в восприимчивом состоянии очень малы. Многие ветроопыляемые растения двудомны (т. е. мужские и женские цветки находятся на разных растениях), например, ивы, однодомны (мужские и женские цветки на одном растении), в частности дубы (см. рис. 18-45), или генетически самонесовместимы (многие злаки).
Рис. 29-34. Цветки злаков обычно закладываются группами. А. Когда такая группа заканчивает развитие, пара сухих жестких брактей (колосковые чешуи) слегка расходится, открывая удлиненный колосок, состоящий из одного — многих цветков (в зависимости от вида злака), прикрепленных к центральной оси. Б. Каждый цветок окружен двумя собственными брактеями — верхней и нижней цветковыми чешуями. Они раздвигаются, открывая внутренние части цветка (В), благодаря набуханию лодикул (мелких округлых телец у основания плодолистика) и широко расходятся к моменту цветения. У тычинок (обычно их три) тонкие тычиночные нити и удлиненные пыльники; рыльца, как правило, вытянутые и перистые, что позволяет эффективно улавливать переносимую ветром пыльцу
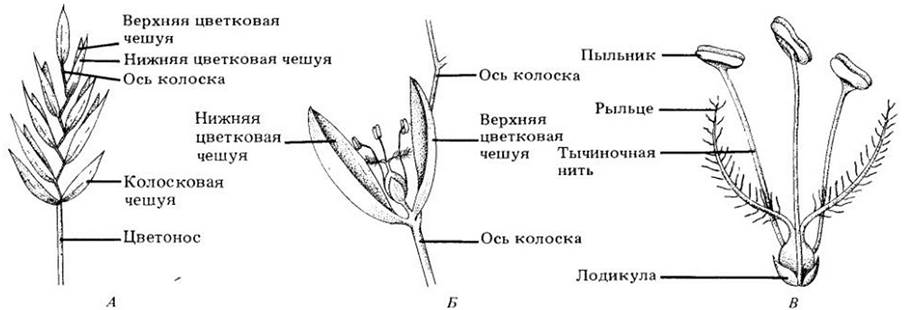
Таким образом, несмотря на случайное перемещение пыльцы, у них существуют механизмы, способствующие высокой степени ауткроссинга.
Из ветроопыляемых цветков тычинки обычно сильно высовываются наружу, так что высыпающаяся пыльца легко подхватывается ветром. У некоторых растений пыльники свешиваются на длинных тычиночных нитях и могут свободно раскачиваться (рис. 29-33 и 29-34). Многочисленные пыльцевые зерна обычно гладкие и мелкие, не слипаются друг с другом, как у насекомоопыляемых видов. Крупные рыльца, как правило, выставлены из цветка и часто разветвлены или имеют перистые выросты, улавливающие переносимую ветром пыльцу. У большинства ветроопыляемых растений в завязи одна семяпочка (а значит, и односемянный плод), поэтому для оплодотворения достаточно попадания на рыльце всего одного пыльцевого зерна. Так, каждый цветок у дуба производит единственный желудь, а у злака только одно зерно, но ветроопыляемые растения с очень мелкими цветками компенсируют это многоцветковостью и(или) многочисленностью соцветий (рис. 29-33, 29-34 и 29-35).
Рис. 29-35. Наиболее обычные для умеренных областей виды деревьев ветроопыляемые. Тычиночные цветки березы бумажной (Betula papyrifera) собраны в свисающие ''сережки'' — гибкие тонкие соцветия длиной несколько сантиметров. Ветер раскачивает их и разносит высыпающуюся зрелую пыльцу

Окраска цветков
Все разнообразие окрасок цветков обеспечивается очень малым набором пигментов. Красная, желтая и оранжевая обусловлена каротиноидами, похожими на те, что присутствуют в листьях. Однако «главные» пигменты цветков — флавоноиды, соединения, в которых два ароматических кольца связаны между собой трехуглеродным фрагментом. Флавоноиды присутствуют, вероятно, у всех покрытосеменных, но лишь изредка встречаются в других группах сосудистых растений, а также у водорослей и животных. В листьях они задерживают ультрафиолетовую радиацию, разрушительно действующую на нуклеиновые кислоты и белки, и обычно избирательно поглощают сине-зеленые и красные лучи, которые важны для фотосинтеза.
Один из крупнейших классов флавоноидов, антоцианы (рис. 29-36), играет ведущую роль в определении окраски цветков. К ним относится большинство красных и синих растительных пигментов. Они растворимы в воде и находятся в вакуолях. Каротиноиды, напротив, жирорастворимы и содержатся в пластидах. Цвет антоцианового пигмента зависит от кислотности клеточного сока в вакуолях; например, цианидин—красный в кислой среде, фиолетовый в нейтральной и синий в щелочной. У некоторых растений окраска цветка меняется после опыления, обычно за счет антоцианов, делающих их менее заметными для насекомых. Кен Пейдж и Томас Уайтмен из Североаризонского университета вблизи Флагстаффа (шт. Аризона) недавно сообщили, что отдельные экземпляры и популяции многолетней травы Ipomopsis aggregata образуют темноокрашенные цветки в начале периода цветения, когда их главными опылителями являются колибри, и светлоокрашенные несколько позже, когда основными опылителями становятся бражники (рис. 29-37). Это, вероятно, зависит от внутриклеточного pH, контролируемого растением.
Рис. 29-36. Три антоциана — основные пигменты, от которых зависит окраска цветка многих покрытосеменных: пеларгонидин (красный), цианидин (фиолетовый) и дельфинидин (синий). Родственные им соединения флавонолы — желтые или кремовые, а каротиноиды — красные, оранжевые или желтые. Бетацианины (беталаины) — красные пигменты, которые встречаются в одной из групп двудольных. Смешение этих разных пигментов при разных pH в клетках образует всю гамму окрасок цветка покрытосеменных. Изменение окраски цветков является «сигналом» для опылителей, сообщающим о том, какие цветки раскрылись недавно, т. е. с большей вероятностью содержат пищу
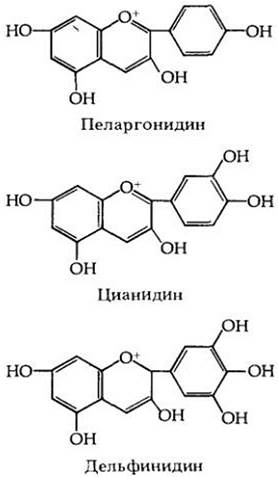
Рис. 29-37. Ipomopsis aggregata из семейства синюховых (Рolemoniaеае) на горе Ферн близ Флагаcпаффа в северной Аризоне цветет с середины июля до сентября. А. В начале и середине цветения отдельные растения и популяции образуют темно-красные цветки, посещаемые колибри. В течение августа популяция колибри в этом регионе постепенно уменьшается и исчезает в начале сентября. Б. В конце сезона растения образуют бледнее окрашенные цветки, более привлекательные для бражников, их главных опылителей в это время. Различия в окраске цветков почти наверняка объясняются изменением клеточного pH, а не образованием новых видов антоцианов
Флавонолы, другая группа флавоноидов, также очень часто содержатся в листьях и цветках. Многие из них вообще или почти бесцветны, но могут придавать цветкам оттенок слоновой кости или белизны.
У всех покрытосеменных характерная пигментация цветка зависит от смешения в разных пропорциях флавоноидов и каротиноидов, клеточного pH, а также структурных, т. е. отражательных, особенностей тканей. Яркая осенняя окраска листьев связана с превращением больших количеств бесцветных флавонолов в антоцианы при разрушении хлорофилла. Цветок калужницы болотной (Caltha palustris) целиком желтый. При этом периферическая часть лепестков, отражающая ультрафиолетовые лучи, окрашена каротиноидами, а поглощающая их центральная часть кажется нам желтой из-за присутствия флавоноида халькона. Для пчел и других насекомых цвет наружной части будет смешанным желто-ультрафиолетовым (так называемый «пчелиный пурпурный»), а неотражающей центральной — чисто желтым (см. рис. 29-23). Чаще всего, хотя и не всегда, способность цветков отражать ультрафиолет связана с присутствием каротиноидов, т. е. ультрафиолетовый узор более присущ желтым цветкам.
У маревых, кактусовых, портулаковых и других представителей порядка Chenopodiales (Centrospermae) красноватый пигмент не антоциановой и даже не флавоноидной природы. Он относится к более сложным ароматическим соединениям, бетацианинам (беталаинам). Именно ими окрашены красные цветки бугенвиллеи (Bougainvillea) и органы свеклы. В растениях перечисленных семейств антоцианов нет, и их биохимические особенности свидетельствуют о близком родстве.
Разнообразие плодов
Плод — это зрелая завязь, иногда с некоторыми другими частями цветка (в последнем случае его называют ложным плодом). Хотя обычно внутри находятся семена, иногда их не образуется, и возникают партенокарпические плоды. Хорошо известный пример последних — бананы.
Обычно различают простые, сборные плоды и соплодия. Это зависит от расположения карпелей. Простые плоды развиваются из одного или нескольких соединенных плодолистиков, а сборные (например, у магнолии, малины и земляники) — из нескольких раздельных карпелей одного гинецея, которые в данном случае дают плодики (сборный плод магнолии показан на рис. 29-12, В). Соплодие состоит из гинецеев нескольких цветков. Например, ананас — соплодие из многих, первоначально раздельных завязей, сидящих на оси соцветия. Разрастаясь, они сдавливают между собой остальные части цветка.
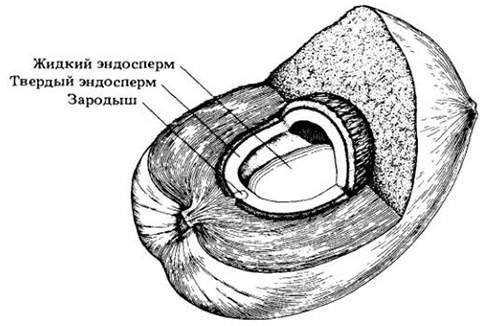
Простые плоды наиболее разнообразны. В зрелом состоянии они могут быть сочными и мясистыми, сухими и одревесневшими, иметь пергаментовидные стенки и т. д. Три главных типа мясистых плодов — ягода, костянка и яблоко. В образовании ягоды, например, томата, финика и винограда, могут участвовать один или несколько, как правило, многосемянных плодолистиков. Внутренний слой ее стенки мясистый. Костянки тоже могут включать один или несколько карпелей, но обычно односемянных, а внутренний слой плода каменистый и, как правило, плотно прирастает к семени. Персики, сливы, оливки и вишни — хорошо известные примеры костянок. Кокосовые орехи тоже костянки, но с волокнистым, а не мясистым наружным слоем (рис. 29-38). Яблоко — специализированный мясистый плод, характерный для одного из подсемейств розовых. Он образуется из нижней завязи, сросшейся с основанием околоцветника, который в основном и дает его мясистую часть. Эндокардии здесь имеет вид жесткой пленки, известной каждому, кто ел яблоки и груши — типичные примеры такого плода.
Рис. 29-38. Плод кокосовой пальмы (Cocos nucífera) является костянкой. Кокосовое молоко — жидкий эндосперм; ко времени прорастания в нем образуются клеточные стенки. Неповрежденный плод легко плавает по морю, за счет чего кокосовые пальмы широко распространились на планете, достигнув самых отдаленных островов. При коммерческой транспортировке с кокосовых «орехов» обычно снимают внешние покровы, так что жители умеренных областей видят снаружи их твердую внутреннюю «скорлупу», непосредственно окружающую семя
Простые сухие плоды делятся на вскрывающиеся (рис. 29- 39 и 29-40) и невскрывающиеся (рис. 29-41). У первых стенка зрелой завязи — перикарпия — растрескивается, высвобождая семена, а у вторых они остаются в плоде и после его опадения с родительского растения. Большинство мясистых плодов невскрывающиеся; они, как правило, содержат одно семя.
Рис. 29-39. Вскрывающиеся плоды. А. Вскрывшиеся листовки ваточника (Asclépios). Б. У некоторых представителей семейства маковых (Papaveraceae), например, у маков (Papaver), семена высеваются из коробочки через отверстия у ее верхушки. В. Для семейства крестоцветных (Brassicaceae) характерен плод, называемый стручком, в котором семена прикреплены к внутренней перегородке, а две прикрывающие ее створки отпадают при созревании

Простые сухие плоды разнообразны. Листовка образована одним плодолистиком; она вскрывается по созревании с одной стороны (примеры — водосбор, ваточник, рис. 29- 39%А). Листовки характерны также для вымершего среднемелового растения Archaeanthus (см. приложение 1 к этой главе) и магнолиевых (см. рис. 29-12, B). Бобовым (Fabасеае) свойствен плод боб, похожий на листовку, но вскрывающийся с обеих сторон (рис. 29-40). В семействе крестоцветных (Вгаssiсасeае) плод стручок. Он образован двумя сросшимися плодолистиками. При созревании две половинки плода разъединяются, а семена остаются прикрепленными к сохраняющейся центральной перегородке (рис. 29-39,5). Наиболее распространенный тип вскрывающегося простого сухого плода — коробочка, образующаяся из сложной верхней или нижней завязи. Семена из нее рассеиваются по-разному. У маковых (Рараverасеае) это часто происходит при ее продольном растрескивании, а у некоторых представителей семейства — через отверстия в верхней части плода (рис. 29-39, Б).
Рис. 29-40. Боб — тип вскрывающегося плода, характерный для бобовых (Faba- сеае, или Leguminosac). Это семейство, включающее около 18 тыс. видов, одно из самых крупных среди цветковых. Многие его представители способны симбиотически фиксировать азот благодаря присутствию на их корнях клубеньковых бактерий из рода Rhizobium (см. с. 145). По этой причине бобовые часто первыми селятся на относительно бесплодных почвах, например, в тропиках, и могут там быстро расти. Семена ряда представителей этого семейства, например, гороха, бобов и чечевицы, являются важными пищевыми продуктами. А. Бобы гороха огородного (Pisum sativum) Б. Бобы «шелковой акации» (Albizziapolyphylla), произрастающей на Мадагаскаре; каждое семя находится в отдельной коучере плода. В. Боб западноафриканского дерева Griffonia simplicifolia. Две его створки разошлись, и внутри видны два семени
Невскрывающиеся простые сухие плоды встречаются во многих семействах (рис. 29-41). Наиболее обычны мелкие односеменные семянки с полостью, в которой свободно лежит семя, прикрепленное только семяножкой. Они типичны для семейств лютиковых (Ranunculасеае) и гречишных (Рolygonасеае). Крылатые семянки, например, у вязов и ясеней, обычно называют крылатками (рис. 29-41, A). Плоды злаков (Роасеае) — зерновки; их семенная кожура плотно прирастает к стенке плода. У сложноцветных (Аsterасеае) сложный семянкоподобный плод образуется из нижней завязи1 (рис. 29-41, Б; см. также рис. 29-43). Желуди и плоды лещины — примеры орехов, похожих на семянки, но имеющих каменистую стенку плода и развивающихся из сложной завязи. Наконец, у зонтичных (Арiасеае) и кленовых (Асегасеае), а также у ряда других неродственных семейств плод схизокарпий, распадающийся при созревании на две или более односемянных частей (рис. 29-42, A).
1В советской литературе он также называется семянкой. — Прим. ред.
Рис. 29-41. Невскрывающиеся плоды. А. Крылатка, характерная для ясеней (Fraxinus) и ильмов (Ulmus), в зрелом состоянии сохраняет единственное семя. Эти плоды распространяются ветром. Б. Семянки лопуха (Arctium) из семейства сложноцветных прицепляются к проходящим животным и таким способом расселяются

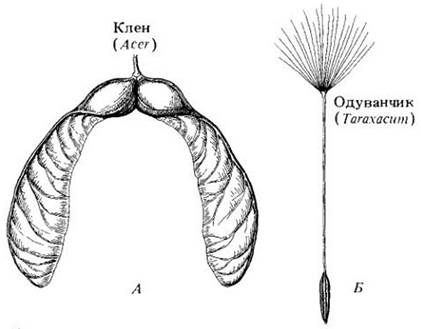
Рис. 29-42. Плоды, распространяемые ветром. А. У кленов (Acer) каждая половинка схизокарпия снабжена длинным крылом. Б. Плоды одуванчика (Taraxacum) и многих других сложноцветных имеют видоизмененную чашечку (nanny с), прирастающую к зрелой семянке, образуя хохолок, что способствует распространению плодов ветром
Распространение плодов и семян
Как цветки можно подразделять в соотвествии с опылителями, так и плоды группируют в зависимости от агентов их распространения.
Плоды и семена, распространяемые ветром
У некоторых растений легкие плоды или семена разносит ветер (рис. 29-39, А; 29-41, А; 29-42), например пылевидные семена орхидных или плоды с крыльями, которые иногда образуются за счет частей околоцветника. Так, у кленов с гинецеем из двух соединенных плодолистиков каждый из них в схизокарпии образует длинное крыло (рис. 29-42, А). При созревании карпели разъединяются и опадают. У многих представителей сложноцветных, например, у одуванчиков, пушистый похожий на парашют паппус способствует парению легкого плода в воздухе (рис. 29-4, Б и 29-43). У некоторых растений крыло или пучок волосков несет не плод, а само семя; у льнянки (Linaria), в частности, семена крылатые, а у кипрея (Epilobium) и ваточника (Asclepias, см. рис. 29-39, А) с хохолком. У ив и тополей (семейство Salicaceae) вся семенная кожура покрыта как бы войлоком. У видов перекати-поля (например, в роде Salsola) все растение (или его часть) гонится по земле ветром, рассыпая при этом семена (рис. 29-44).
Рис. 29-43. Всем известные мелкие невскрывающиеся плоды одуванчика представляют собой семянки, распространяющиеся благодаря видоизмененной в хохолок чашечке (паппусу). На фотографии показаны корзинки растения из рода Agoseris, близкородственного одуванчикам, с плодами
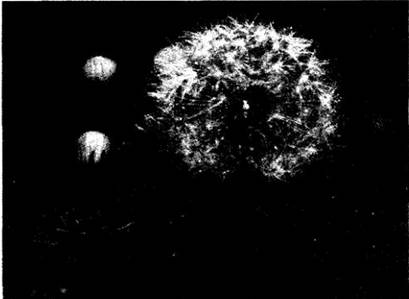
Рис. 29-44. У растений типа перекати- поле (на рисунке — из рода Salsola) надземная часть отрывается от корня и гонится ветром по открытой местности. Семена при этом высыпаются. Родина таких видов Евразия, но они широко распространились как сорняки в Северной Америке и других частях света

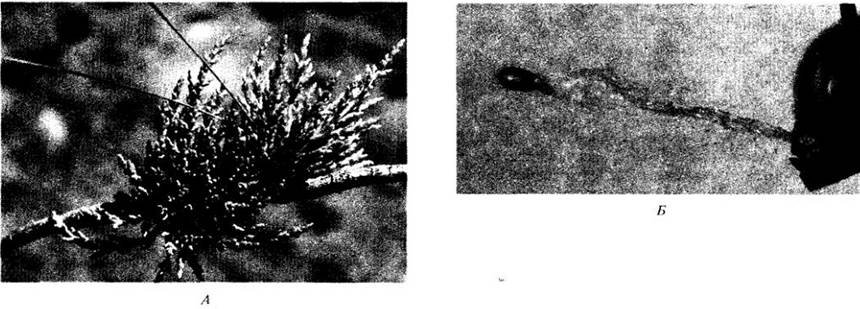
Некоторые растения «выстреливают» свои семена. Так, у недотроги (Impatiens) их выбрасывают на некоторое расстояние внезапно разделяющиеся створки коробочек. У гамма- мелиса (Hammamelis), когда плод высыхает, эндокарпий сжимается, разбрасывая семена с такой силой, что они могут пролетать до 15 м. Другой пример такого рода показан на рис. 29-45. В противоположность этим активным способам распространения семена и плоды многих растений просто опадают на землю и затем перемещаются более или менее пассивно, в частности периодически действующими агентами, например, паводками.
Рис. 29-45. Arceuthobium — паразитическое двудольное, являющееся одной из важнейших причин снижения продуктивности лесов запада США. А. Растение из этого рода на ветке сосны в Калифорнии. Б. Выбрасывание семени. В плоде создается очень высокое гидростатическое давление, и семена выстреливаются в сторону от него на расстояние до 15 м с начальной скоростью около 100 км/ч. Это один из способов переноса семян с дерева на дерево, хотя они клейкие и могут распространяться гораздо дальше, прилипая к ногам или перьям птиц
Плоды и семена, распространяемые водой
Плоды и семена многих видов, особенно произрастающих в водоемах или вблизи них, способны плавать. Это объясняется наличием в них либо особых воздушных камер, либо рыхлых тканей с заполненными воздухом межклетниками. Некоторые плоды специально адаптированы к распространению океаническими течениями, например, у кокосовой пальмы (см. рис. 29-38); именно поэтому она быстро заселяет почти все относительно недавно возникшие в Тихом океане атоллы. Дождь тоже является обычным агентом распространения плодов и семян; он особенно важен для растений, растущих на склонах гор и холмов.
Плоды и семена, распространяемые животньми
Сладкие и часто яркоокрашенные мясистые плоды — несомненно, свидетельство коэволюции животных и растений. В большинстве случаев те из них, у которых значительная часть перикарпия мясистая (например, слива, малина, кизил, виноград), поедаются позвоночными (обычно млекопитающими или птицами), и тогда содержащиеся в них семена, проходя пищеварительный тракт или отрыгиваясь животным, оказываются на большом расстоянии от места, где они были проглочены (рис. 29-46). Иногда неполное переваривание, размягчающее семенную кожуру, способствует прорастанию семян.
Созревая, мясистые плоды претерпевают ряд характерных изменений под действием гормона этилена (см. гл. 24). В них увеличивается содержание сахара, ткани размягчаются за счет разрушения пектиновых веществ, а зеленоватый, как у листьев, незаметный цвет сменяется ярко-красным (рис. 29-46, А), желтым, синим или черным. Семена некоторых растений, особенно в тропиках, имеют яркоокрашенные мясистые придатки, ариллусы. Их сочность и окраска способствуют, как и у плодов, распространению семян позвоночными. Ариллусы тисса (Taxus; см. рис. 18-26) не гомологичны одноименным структурам покрытосеменных, т. е. имеют иное происхождение, но выполняют ту же функцию.
Рис. 29-46. Семена мясистых плодов обычно распространяются плодоядными позвоночными. При этом семена либо отрыгиваются, либо выходят наружу с фекалиями. А. Земляника (Fragaria) — пример сборного плода с семянками на поверхности мясистого цветоложа. В незрелом виде эти плоды, как и многие другие, распространяемые птицами и млекопитающими, зеленые. Они краснеют, когда семена созревают и становятся готовыми для расселения. Б. Ягоды многих кактусов, например, опунции (Opuncia) на юге Мексики, становятся хорошо заметными при созревании. В. Ягоды жимолости Lonicera hispidula; у этого вида плоды образуются из нижней завязи и включают слившиеся части наружных мутовок цветка, показанного на рис. 29-13, В

Часто зеленая или маскирующая окраска незрелых плодов в определенной степени защищает их от преждевременного поедания животными. При этом они могут быть неприятными на вкус, как очень кислые незрелые вишни (Prunus), что также отталкивает животных. Изменением окраски при созревании плодов растение «сигнализирует» об их съедобности, т. е. о том, что семена созрели и готовы к распространению (рис. 29-46). В это время неслучайно преобладает красный цвет. Благодаря ему плоды незаметны для насекомых — по-видимому, сливаются для них с зеленым фоном листьев. Эти животные слишком малы, чтобы эффективно распространять крупные семена мясистых плодов, и привлекать их растению не выгодно. В то же время красные плоды хорошо заметны для позвоночных, которые, поедая их, переносят зрелые семена на крупные расстояния.
Плоды или семена многих покрытосеменных распространяются, прикрепляясь к шерсти или перьям животных (рис. 29-47; 29-41,5), и снабжены для этого крючками, бородками, шипами, щетинками или клейкими покровами.
Рис. 29-47. Плоды африканского растения Harpagophytum, представителя семейства кунжутовых (Pedaliaceae), снабжены цепкими крючками, с помощью которых прицепляются к шерсти на ногах крупных млекопитающих и переносятся ими с места на место

Биохимическая коэволюция
Еще один важный фактор эволюции покрытосеменных связан так называемыми вторичными метаболитами растений, которые раньше считались отходами их жизнедеятельности. Сюда относится множество соединений из разных классов — алкалоиды, хиноны, эфирные масла (включая терпеноиды), гликозиды (включая цианогенные вещества и сапонины), флавоноиды и даже рафиды, т. е. игольчатые кристаллы оксалата кальция. Некоторые из них характерны для целых семейств или групп семейств цветковых (рис. 29-48).
Рис. 29-48. Вторичные растительные метаболиты: синигрин из горчицы черной (Brassica nigra); калактин, сердечный гликозид из ваточника Asclépios curassavica; никотин табака Nicotiana tabaccum, представителя пасленовых; кофеин кофейного дерева (Coffea arabica) из семейства мареновых; теобромин, главный алкалоид кофе, чая (Thea sinensis) и какао (Theobroma cacao). Никотин, кофеин и теобромин — алкалоиды, представители разнообразного класса азотсодержащих циклических соединений, физиологически активных для позвоночных

В природе эти вещества, по-видимому, играют главную роль в ограничении съедобности содержащих их растений, заставляя многих животных их избегать (рис. 29-49). Если растения какого-то семейства содержат определенные вторичные метаболиты, поедать их могут только насекомые из определенных семейств. Например, для крестоцветных (Вrassicасеае) характерно присутствие гликозидов горчичного масла и ферментов, разрушающих их с выделением продуктов с едким запахом, характерным для капусты, хрена и горчицы. Большинство насекомых-фитофагов избегает растения этого семейства, несмотря на потребность в пище. Однако некоторые клопы, жуки и личинки чешуекрылых питаются только листьями крестоцветных, например, гусеницы многих видов бабочек из подсемейства Pierinae (сюда относятся так называемые капустницы, репницы и др.). Одни и те же химические вещества, отпугивая основную массу насекомых-фитофагов, стимулируют пищевую активность видов с узкой пищевой специализацией. Например, если личинок некоторых чешуекрылых, питающихся капустой, поместить на агар или фильтровальную бумагу, смоченные капустным соком, они вытягивают свой ротовой аппарат и совершают характерные «пищевые» движения.
Рис. 29-49. Toxicodendron radicans вырабатывает вторичный метаболит 3- пентадекандиенилкатехол, вызывающий зудящую сыпь на коже многих людей. Способность к синтезу этого спирта возникла у него, вероятно, в условиях отбора, производимого фитофагами. К счастью, это растение легко узнать по его характерным сложным листьям с тремя листочками

Ясно, что способность производить эти химические вещества и накапливать их в тканях — важное эволюционное приобретение соответствующих растений, обеспечивающее им биохимическую защиту от большинства фитофагов. Для этого, безусловно, служат гликозиды горчичного масла у крестоцветных. Что касается фитофагов, то для любой группы насекомых, устойчивых к этим ядам или способных разрушать их, защищенные таким способом растения — обильный в связи с непоедаемостью другими насекомыми источник питания. Так, быстрое эволюционное развитие подсемейств; Pierinae, вероятно, началось только после приобретение предковыми формами способности питаться крестоцветными, расщепляя их токсины.
Насекомые-фитофаги, рацион которых ограничен узким кругом растений с определенными вторичными метаболита ми, часто имеют яркую окраску, предупреждая тем самым хищников о присутствии в своем теле ядовитых веществ. Так, среди насекомых, питающихся в дневное время ваточником, — ярко-зеленые жуки-листоеды, ярко-красные жуки дровосеки и клопы, черные бабочки данаиды. Представители семейства листовневых (Asclepiadaceae). к которык относится это растение, богаты алкалоидами и сердечным! гликозидами, т. е. ядами, сильно действующими на позвоночных, потенциальных врагов этих насекомых. У птицы, проглотившей данаиду, начнется сильное расстройство желудка с рвотой, и такой оранжево-черный узор, как на крылья: этой бабочки, хищник в дальнейшем будет избегать. У других насекомых, например, у американской бабочки Basila rchia archippus, выработалась сходная окраска крыльев, т. е они спасаются от врага, имитируя внешность ядовито! данаиды. Это явление, называемое мимикрией, в конечном счете оказывается зависящим от химических защитны: веществ. Различные наркотики и психотомиметические пре параты, например, биологически активные соединения конопли (Cannabis sativa) и мака снотворного (Papaver somniferum), также являются растительными вторичными метаболитами, роль которых в природе заключается в отпугивании фитофагов (рис. 29-50).
Рис. 29-50. Некоторые растения, вырабатывающие галлюциногенные и лекарственные вещества. А. Мескалин из кактуса лофофора Вильямса (Lophophora williamsii) применяется на обрядовых церемониях многими группами индейцев северной Мексики и юга-запада США. Б. Тетрагидроканнабинол — основное биологически активное вещество конопли (Cannabissativa). В. Хинин, ценное лекарство для лечения и предупреждения малярии, добывается из тропических деревьев и кустарников рода Cinchona. Г. Кокаин, наркотик, которым недавно сильно злоупотребляли, получают из кокаинового куста (Erythroxylon coca), выращиваемого на северо-западе Южной Америки. На фотографии изображена перуанка, собирающая листья этого растения. Вторичные метаболиты, вероятно, защищают его от насекомых-фитофагов, но физиологически активны также в отношении позвоночных, включая человека

Известны и более сложные защитные системы. Когда листья томата или картофеля повреждены, например, колорадским жуком, концентрация ингибиторов — протеиназ, блокирующих пищеварительные ферменты насекомого в контактирующих с воздухом тканях растений — быстро возрастает. Другие растения вырабатывают молекулы, близкие по структуре к гормонам насекомых или других фитофагов, и таким способом нарушают нормальный рост и развитие животных. Большой интерес для человека представляет сложное по строению вещество диосгенин, получаемое главным образом из дикого ямса, произрастающего в Мексике, и в меньших количествах также из индийского и китайского видов ямса. В химическом отношении диосгенин очень близок к известному химическому соединению 16-дегидропрег- ненолону (16D) — главному действующему началу многих пероральных противозачаточных средств. Дикий ямс раньше был основным сырьем для его производства. К сожалению,
он растет очень медленно и его природные запасы скоро могут иссякнуть. Исследователи в СССР, Эквадоре и других странах изучали возможности возделывания определенных видов паслена (Solanum), содержащих соласодин — вещество, которое легко превратить в 16D. Некоторые из этих растений уже выращиваются в коммерческих целях.
Выше уже говорилось, что в ходе коэволюции систем опыления каждый их тип появлялся не один, а, как правило, несколько раз. Возникшее в результате разнообразие способов переноса пыльцы чрезвычайно сильно расширило возможности дифференциации цветковых. Однако в случае биохимических особенностей этапы эволюции прослеживаются очень четко. Можно охарактеризовать с этой точки зрения целые семейства растений, показав их связь с главными группами насекомых-фитофагов. Не исключено, что эти биохимические связи играли ключевую роль в первоначальном успехе покрытосеменных.
Заключение
Древнейшие ископаемые остатки покрытосеменных, доминирующей в настоящее время группы сосудистых растений, имеют возраст около 125 млн. лет (начало мелового периода). Примерно 80 — 90 млн. лет назад (поздний мел) они стали господствовать на всем земном шаре. Среди растительных остатков в отложениях этого времени можно определить многие современные семейства покрытосеменных и даже отдельные современные роды. Не исключено, что пыльцу покрытосеменных, существовавших более 125 млн. лет назад, мы нс способны отличить от пыльцы голосеменных или спор папоротников, поэтому трудно с уверенностью говорить о более раннем присутствии цветковых на планете, но, безусловно, эта группа несколько старше.
Покрытосеменные могли возникнуть на семиаридных возвышенностях и в сухих внутренних бассейнах запада Гондваны — суперконтинента, давшего начало Южной Америке и Африке. К тому времени, когда эти континенты полностью разделились (около 90 млн. лет назад), их климат существенно изменился, и покрытосеменные начали захватывать мировое господство. В числе возможных причин их успеха можно назвать различные адаптации, повышающие засухоустойчивость, а также возникновение эффективных и высокоспециализированных систем опыления.
Важнейший признак покрытосеменных — цветок, сыгравший важную роль в их эволюции. Плодолистик — продольно сложенная листоподобная структура, защищающая семязачатки (содержащие мегаспорангии), дифференцировалась на базальную расширенную часть — завязь, удлиненный столбик и улавливающее пыльцу рыльце. Тычинки также возникли от листоподобных образований или тонких разветвленных систем с терминальными спорангиями. Чашелистики — специализированные листья, защищающие цветок в бутоне. Лепестки у некоторых покрытосеменных — стерилизовавшиеся тычинки, принявшие на себя функцию привлечения насекомых, однако у других они произошли от чашелистиков. Спиральное расположение, многочисленность и раздельность частей цветка примитивных покрытосеменных сменились у большинства современных форм мутовчатым расположением вполне определенного их числа и слиянием внутри одной или даже между разными мутовками.
Примеры специализированных семейств — Asteraceae (сложноцветные), у которых многочисленные высокоспециализированные цветки собраны в соцветие корзинку, функционирующее как единая, привлекающая насекомых структура, и Orchidaceae (орхидные) с причудливыми очертаниями частей билатерально-симметричного цветка, имеющего наиболее специализированную систему опыления.
Опыление насекомыми — одна из важнейших черт покрытосеменных; первыми его агентами могли быть жуки или подобные им животные. Смыкание краев плодолистика, вероятно, связано с защитой семязачатков от поедания опылителями. Более специализированные группы насекомых возникли позднее, коэволюционировав вместе с покрытосеменными; осы, мухи, дневные и ночные бабочки оставили свой след в морфологии цветка некоторых групп. Однако самые специализированные и постоянные «посетители» цветка — пчелы; они оказали, вероятно, наибольшее влияние на его эволюцию. Каждая группа посещающих цветок животных связана с особым набором его признаков, воздействующих на зрение и обоняние. Некоторые покрытосеменные стали ветроопыляемыми и рассеивают большое количество мелкой, не слипающейся пыльцы; у них хорошо развитое, часто перистое рыльце, эффективно улавливающее ее из воздуха.
У растений, которые регулярно посещаются и опыляются животными, нуждающимися в энергетически богатой пище (например, колибри), цветки должны производить большое количество нектара, при этом скрывая и защищая его источники от других возможных посетителей с меньшими энергетическими потребностями, т. е. способных удовлетвориться нектаром одного цветка (или цветков одного растения) и малоэффективных в смысле перекрестного опыления. Опыление ветром не так надежно; при этом отдельные растения должны расти близко друг к другу крупными группами, тогда как насекомые, птицы и летучие мыши могут переносить пыльцу даже между цветками, разделенными большим расстоянием.
Плоды так же разнообразны, как и цветки, из которых они образуются. Их можно классифицировать как по морфологии и анатомии, так и по способам распространения. Плоды — это зрелые завязи, иногда с сохраняющимися при них другими частями цветка (ложные плоды). Простые плоды образуются из одного или нескольких соединенных плодолистиков, сборные — из свободных плодолистиков одного цветка, а соплодия — из плодолистиков нескольких (иногда многих) цветков. Вскрывающиеся плоды рассеивают семена растрескиваясь; невскрывающиеся — другим способом.
Переносимые ветром плоды или семена легкие, часто крылатые или с пучками волосков, что способствует их распространению. Некоторые плоды разбрасывают семена взрываясь. Многие плоды или семена разносятся водой; они должны быть плавучими и иметь водостойкие покровы. Другие распространяются животными, в основном позвоночными, и отличаются мясистым околоплодником, привлекающим своим вкусом и часто броским видом фитофагов. Некоторые плоды распространяются, прикрепляясь к покровам млекопитающих или к перьям птиц.
Третья причина успеха покрытосеменных и увеличения их разнообразия — биохимическая коэволюция. В некоторых группах цветковых выработалась способность образовывать вторичные метаболиты (например, алкалоиды), защищающие их от большинства фитофагов. Однако некоторые растительноядные формы (обычно с узкой пищевой специализацией) способны питаться этими растениями и постоянно сопутствуют им. При этом конкуренция с другими фитофагами исключена благодаря токсичности растений. Таким образом, коэволюция могла происходить ступенчато, и представляется вероятным, что ранние цветковые были защищены ядовитыми для животных веществами.
Приложение 1. Древние покрытосеменные и их цветки
Archaeanthus linnenbergeri, единственный известный представитель вымершего семейства покрытосеменных, произрастал в середине мелового периода (90 — 95 млн. лет назад) на месте центрального Канзаса. В простиравшихся здесь субтропических прибрежных равнинах было много динозавров. Плотные цветки Archaeanthus имели удлиненную ось, несущую 100 — 130 спирально расположенных листовок с 10 — 18 семенами в каждой. Околоцветник состоял из трех наружных частей и 6— 9 внутренних; многочисленные тычинки шли по спирали. Вероятно, это было листопадное деревце или кустарник, морфологией цветка в общих чертах напоминающий современных представителей семейств, родственных магнолиевым (см. рис. 29-11). А — отпечаток репродуктивной оси; Б — ископаемый лист; В — реконструкция ветки с цветком; Г — реконструкция ветки с плодами. Реконструкции сделаны на основе работ Д. Дилкера из Университета Индианы; рисунки выполнены М. Роном. Тщательные исследования Дилкера и его учеников много дали для понимания природы ранних покрытосеменных и их цветка.

Приложение 2. Генетическая самонесовместимость
Рекомбинация генов различных особей в ходе полового воспроизводства создает разнообразие естественных популяций, придаст им способность адаптироваться к изменениям окружающей среды путем постепенных эволюционных преобразований. Самоопыляющиеся растения имеют в этом смысле меньше возможностей. На заре истории цветковых во многих их семействах выработались механизмы, делающие перекрестное опыление обязательным даже при обоеполых цветках или однодомности.
Среди современных видов известны два основных механизма, стимулирующие этот процесс. Чаще всего (например, у таких важных в хозяйственном отношении растений, как злаки и бобовые) судьба пыльцевого зерна определяется его собственным (гаплоидным) генотипом. Если в локусе несовместимости оно несет такой же ген, что и в соответствующем локусе у диплоидного рыльца и столбика, путь пыльцевой трубке закрывается. Если ген этого локуса у пыльцевого зерна не такой, как в ткани рыльца, оно нормально прорастает.
В иного рода системе, обнаруженной в семействах крестоцветных (Вrassicaceae) и сложноцветных (Аsteraceae), будущее пыльцы определяется генами произведшего ее растения, а нс отдельного зерна, т. е. соответствием диплоидных тканей обоих родителей. В обоих случаях возможность оплодотворения определяется сочетанием генов локуса несовместимости.
Хотя еще много предстоит узнать о физиологии этих механизмов, уже ясно, что они зависят от реакций «узнавания» между специфическими белками пыльцевых зерен и тканей рыльца или столбика. У злаков эта реакция часто происходит на поверхности рыльца. На фотографии, сделанной с помощью сканирующего электронного микроскопа (А), показана часть рыльца ежи сборной (Dactylis glomerata). Оно покрыто многочисленными сосочками, каждый из которых способен уловить несколько пыльцевых зерен. На рис. Б можно видеть разрез стенки такого выроста у другого злака — ржи (Secale cereale) — в просвечивающем электронном микроскопе. Над кутикулой находятся еще два слоя, внутренний — из слизистых пектиновых веществ и наружный — белковый. Известно, что несовместимость проявляется, когда пыльцевая трубка соприкасается с наружным слоем или вскоре после этого. На двух других микрофотографиях (В, Г) у сделанных с помощью флуоресцентного микроскопа, показаны рыльца лисохвоста (Alopecurus pratensis), окрашенные флюоресцентным красителем для выявления полисахарида клеточной стенки, называемого каллозой. На рис. В опыление совместимое (видно, как, проникнув в рыльце, трубка растет в направлении завязи), на рис. Г — несовместимое: после соприкосновения конца трубки с белковым слоем ее рост остановился, а внутренняя часть заполнилась каллозой, что означает «отторжение» пыльцевого зерна.