СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IX. ЭКОЛОГИЯ
ГЛАВА 31. ДИНАМИКА СООБЩЕСТВ И ЭКОСИСТЕМ
Экология — это наука о взаимодействиях организмов друг с другом и с окружающей средой. Она пытается объяснить, почему те или иные растения и животные встречаются только в определенном регионе, почему так много организмов одного типа и так мало другого, какие изменения могут вызвать их взаимодействия на данной территории и как функционируют экосистемы, особенно с точки зрения образования и использования органических соединений и круговорота элементов.
Для последующих рассуждений будут полезны некоторые определения. Сообщество состоит из всех растений, животных и других организмов, живущих на определенной площади.Экосистема включает не только эти живые организмы (биотические факторы), но также и неживые (физические) элементы окружающей среды, с которыми они взаимодействуют. Биомы — это крупные территориальные комплексы сообществ, характеризующиеся особыми растительностью и климатом, например, пустыни или степи. Важнейшие биомы описаны в гл. 32.
Взаимодействия между организмами
Ни один организм в сообществе (неважно, в лесу, на пастбище, в пруду или на коралловом рифе) не существует изолированно от своего окружения. Он взаимодействует либо с другими живыми существами, либо с факторами окружающей среды. В этой главе будут обсуждены три главных типа взаимодействий между организмами: мутуализм, конкуренция и отношения растений с растительноядными животными (фитофагами).
Мутуализм
Мутуализм — это биологическое взаимодействие, способствующее росту и выживанию обоих видов-партнеров, т. е. одна из форм симбиоза. В природе ни один из таких видов не может жить без другого. Классический пример такого симбиоза — лишайники — уже обсуждался в гл. 13. Другой пример — отношения между бобовыми и азотфиксирующими бактериями в клубеньках на их корнях (см. гл. 26). Мутуализмом являются и некоторые тесные взаимосвязи опылителей с опыляемыми растениями, обсуждавшиеся в гл. 29, например, чешуекрылого Tegeticula yucasella и юкки.
Один из наиболее интересных и экологически важных примеров мутуализма — взаимодействия между грибами и сосудистыми растениями. Как уже говорилось в гл. 13 и 26, корни большинства этих растений образуют с грибами сложные структуры, называемые микоризой. Без нее нормальный рост растений был бы невозможен. Микориза, по-видимому, сыграла решающую роль при заселении ими суши.
Чем больше мы узнаем о микоризных связях, тем яснее становится их важность для сосудистых растений. У многих их видов немикоризные особи в природе встречаются редко, даже если их рост и возможен без грибов при тщательном подборе условий произрастания. Большинство сосудистых растений — «двойные» организмы в том же смысле, что и лишайники, хотя эта двойственность как правило незаметна по их надземной части. По словам почвоведа, из Висконсинского университета С. Уайлда, «дерево, извлеченное из почвы, — это только часть целого растения, хирургически отделенная от его... поглощающего и пищеварительного органа». У большинства растений микоризные грибы играют жизненно важную роль в усвоении фосфора и других необходимых питательных веществ.
Грибы, образующие микоризу с большинством растений, относятся к зигомицетам. Как детально обсуждалось в гл. 13, такой се тип называют эндомикоризой; она характерна для большинства трав, кустарников и деревьев. Некоторые группы хвойных и двудольных — в основном деревья — образуют микоризу с базидиомицетами, а также с некоторыми аскомицетами. В этом случае речь идет об эктомикоризе. Иногда она очень специфична: один вид гриба взаимодействует только с определенным видом сосудистого растения или с группой родственных видов. Известно, например, что базидиомицет Boletus elegans связан только с лиственницей (Larix) из хвойных. Другие грибы, например, Сепососсит geophilum,образуют микоризу с лесными породами более десятка родов. Эктомикориза особенно характерна для относительно бедных видами сообществ деревьев, живущих в высоких широтах Северного полушария или в высокогорьях.
Наиболее сложные примеры мутуализма встречаются в тропиках, где разнообразие организмов гораздо выше, чем в умеренных областях. Так, в тропиках и субтропиках широко распространены акации (деревья и кустарники рода Acacia). Взаимосвязи между определенными видами акаций на равнинах Мексики и Центральной Америки и муравьями, обитающими в их шипах, — замечательный пример сложных взаимодействий между животными и растениями. Особенно наглядно они прослеживаются для муравьев рода Pseudomyrmех (рис. 31-2).
Рис. 31-2. Муравьи и акации. А. Основание черешка одного из сложных листьев с нектарниками. Б. Тельца Белта на кончиках листочков A. collinsii. В. Муравьи-рабочие (Pseudomyrmex ferruginеа), нападающие на усики лианы, зацепившиеся за A. cornígera. Получая всю свою пищу от акации, муравьи в свою очередь устраняют все растения, вступающие в контакт с этим деревом, и убивают большую часть других насекомых, которые делают попытки питаться на нем. Г. Отдельный экземпляр A. cornígera, возвышающийся над густой вторичной растительностью на тропической равнине Мексики. Если бы на этом растении не поселились муравьи, оно, вероятно, погибло бы еще на стадии всхода, затененное другими растениями или съеденное насекомыми


У этих акации в основании каждого листа находится пара вздутых шипов длиной более 2 см, на черешках расположены нектарники, а на кончиках листочков — мелкие питательные органы, называемые тельцами Белта. Муравьи живут внутри полых шипов, питаясь сахарами из нектарников и тельцами Белта, содержащими жиры и белки. Акации растут чрезвычайно быстро и особенно характерны для нарушенных территорий, где конкуренция между быстрорастущими растениями-колонизаторами часто очень интенсивна.
Томас Белт впервые описал взаимоотношения между Pseudomyrmex и этими деревьями в своей книге «Натуралист в Никарагуа» («The Naturalist in Nicaragua»), вышедшей в 1874 г. После его наблюдений велись долгие споры о том, действительно ли присутствие муравьев приносит акациям пользу. Этот вопрос был окончательно решен в 1964 г. Д. Джэн- зеном, в то время студентом Калифорнийского университета в Беркли, а ныне профессором Пенсильванского университета. Он обнаружил, что муравьи-рабочие, кишащие на поверхности растения, нападают на животных, любых размеров, которые так или иначе прикасаются к нему, таким образом одновременно защищая акацию от фитофагов и сохраняя жилье и пищу для себя и своих собратьев. Более того, если занятой ими акации касаются ветви другого растения, муравьи обдирают с них кору. В результате эти ветви отмирают, что обеспечивает доступ акации, окруженной быстро растущей тропической растительностью, к свету.
Когда Джэнзен удалял муравьев (отравляя их или отламывая населенные ими части растения) или наблюдал изредка встречавшиеся акации без этих животных, он обнаружил, что рост деревьев крайне замедлялся и обычно они погибали через несколько месяцев в результате повреждения насекомыми или затенения другими видами. И напротив, акации с муравьями росли очень быстро, вскоре достигая 6 м и более в высоту и затеняя более низкие растения. Эти муравьи строят свои гнезда только на деревьях определенного вида и полностью зависят от их нектарников и телец Белта как источников питания. Таким образом, система муравей — акация — такой же двойной биологический комплекс, как, скажем, лишайник. Один элемент не может выжить без другого в сообществе, где возникло это явление.
Существует много других типов связывающих организмы мутуалистических отношений. Например, деревья в лесу (а также и травы) часто сращены своими корнями. В результате питательные вещества передаются от одного растения к другому сложным и совершенно неожиданным путем, и выживание какого-то вида в определенной местности буквально зависит от присутствия другого, с которым он образует такую связь. Пни деревьев могут жить неопределенно долго, хотя и лишены фотосинтезирующих органов, потому что сращены корнями с другими особями и могут получать от них питательные вещества. Некоторые болезни, например, дубовый вилт на Среднем Западе и востоке США, также могут передаваться через такие корневые «прививки».
Конкуренция
В отличие от животных растения получают энергию единственным путем — за счет фотосинтеза. Следовательно, конкуренция между ними проявляется главным образом в «борьбе за свет», и особи, обычно растущие в тени других, выработали различные механизмы, позволяющие фотосинтезировать при низкой освещенности. Хотя важную роль играет также конкуренция за воду и питательные вещества, конкуренция за свет у растений гораздо сильнее. Различия в высоте, расположении листьев, форме кроны — основные факторы, позволяющие конкретным типам растений адаптироваться к различным местообитаниям в пределах одного сообщества, будь то низкотравный луг или высокоствольный лес. У животных конкуренция более специфична. У растений те ее типы, которые мы обсуждаем здесь, могут возникать и между видами, случайно оказавшимися рядом. В большинстве случаев всходы гораздо чувствительнее к конкуренции, чем развитые особи того же вида.
В экспериментальных условиях, когда два вида сосуществуют достаточно долго в однородной среде, один из них всегда погибает (элиминируется). На основе таких наблюдений и теоретических расчетов сложилось мнение, что в природе два вида нс могут сосуществовать бесконечно в одном и том же местообитании, используя одинаковые ресурсы одинаковым способом. Это упрощенное выражение того, что Гарретт Хардин назвал принципом конкурентного исключения. Если два вида растут вместе, потребляя одни и те же незаменимые для них ресурсы, представленные в ограниченном количестве, то особи одного или обоих видов будут меньшего размера или малочисленнее, чем в том случае, если бы виды росли порознь. Когда среда достаточно сложна (а так и бывает в природе), различные организмы могут использовать ее разными способами, в сущности подразделяя местообитания. Тогда они могут сосуществовать неограниченно долго.
Например, на болоте сфагновые мхи часто образуют сплошные ковры из нескольких произрастающих вместе видов. Как им удается сосуществовать? Если проанализировать ситуацию более тщательно, здесь можно обнаружить полуводные виды, растущие по дну наиболее сырых понижений, мхи, занимающие более сухие места по склонам кочек, в образовании которых они участвуют, и виды, выдерживающие только наиболее сухие условия на вершинах кочек, где они в конце концов замещаются одним или несколькими видами цветковых растений. Следовательно, хотя все эти мхи сосуществуют в том смысле, что присутствуют в одном и том же болоте, они фактически занимают различные микроэкотопы и постоянно сменяют друг друга в соответствии с изменениями микросреды.
Если численность популяций сосуществующих видов поддерживается на низком уровне, они могут и не элиминировать друг друга. Это можно проследить на примере Англии, где сильная эпидемия вирусного заболевания миксоматоза катастрофически сократила в нашем веке популяцию кроликов. Прежде злаки на меловых почвах буквально «выстригались» этими млекопитающими, и в таких местообитаниях могли расти многие виды двудольных. После сокращения численности кроликов злаковый покров здесь стал выше и гуще и многие прежде обильные виды разнотравья сделались редкими (рис. 31-3). Сходный эффект часто можно наблюдать, сравнивая выпасаемые и невыпасаемые луга, а также очень часто в районах, пострадавших от природных бедствий, например, от ураганов. По этой же причине можно ожидать большего разнообразия видов вдоль полосы морского прибоя (где нарушения происходят постоянно), чем в более стабильных местообитаниях.
Рис. 31-3. Национальный заповедник «Лаллингтон-Хит» в Восточном Суссексе, Англия. Меловые луга до (А) и после (Б) гибели кроликов в результате миксоматоза. Первая фотография сделана в 1954 г., вторая — в 1978 г. Чтобы восстановить разнообразие травянистых растений, была разработана программа выпаса овец и лошадей на большей части этого заповедника площадью 62 га

Большинство конкурентных ситуаций очень сложно и затрагивает как внутри-, так и межвидовые взаимоотношения. Существуют различные способы выражения относительной конкурентоспособности двух растущих вместе видов. Например, на рис. 31-4 показана зависимость сухой массы урожая с 1 га от густоты посевов кукурузы (Zea mays). По мерю увеличения густоты этот урожай (включая побеги и початки) возрастает. Однако, когда на 1 м2 находится более 7 растений, масса побегов без початков увеличивается быстрее, чем общая масса растений, за счет заметного снижения массы початков.
Рис. 31-4. Влияние густоты растений на урожай кукурузы (Zea mays), измеренный тремя различными способами. Если ставится задача получить максимальную сухую массу початков, оптимальное число растений на 1 м2 должно быть между 7 и 8
Клоновое размножение, важное для многих растений, иногда затрудняет определение границ генетически идентичных особей в природе. Это в свою очередь может препятствовать оценке их конкурентных отношений. Генетически идентичные особи могут широко распространяться в далеко отстоящих друг от друга, но сходных по условиям экотопах внутри сложно организованной среды. Это характерно не только для корневищных многолетников (например, многих злаков и осок), но и для одуванчиков и других видов, у которых семена образуются бесполым путем и содержат зародыши, генетически идентичные родительским особям, а также для растений типа клевера ползучего (Trifolium repens), у которых от исходного экземпляра просто отделяются дочерние особи.
При некоторых видах конкуренции один или оба конкурирующих организма вырабатывают химические вещества, которые подавляют рост особей либо своего вида, что приводит к увеличению расстояния между ними, либо другого вида. Например, гриб Pénicillium chrysogenum, растущий на таком органическом субстрате, как семена, образует значительное количество пенициллина. Этот антибиотик подавляет рост бактерий, непосредственно конкурирующих с грибом за питательные вещества. Однако бактерия Bacillus сегеus,вырабатывающая ферменты пенициллиназы, которые разрушают пенициллин, часто вытесняет этот гриб.
Аналогичные взаимоотношения среди растений часто называют аллелопатией. Например, на калифорнийском побережье между кустарниками и злаковыми сообществами обычно находится своего рода мертвая зона (рис. 31-5). Именно в ней наиболее активно добывают пищу кролики, грызуны и птицы, находящие в кустах убежище. Если проникновению сюда животных препятствует проволочная сетка, на этом обычно пустом пространстве буйно разрастаются однолетние травы (рис. 31-6). Кроме того, некоторые кустарники, например, шалфей Salvia leucophylla, вырабатывают летучие терпены, препятствующие выживанию в непосредственной близости от них всходов многих видов. При этом всходы не могут поселиться в «мертвой» зоне, даже если заграждение не дает проникнуть в нее травоядным животным, но часто растут на границе между ней и ненарушенным злаковником, где концентрация токсинов, вырабатываемых шалфеем, уже невелика, а травостой еще не слишком густой.
Рис. 31-5. Кустарниковый шалфей Salvia leucophylla вырабатывает терпены, которые, испаряясь, распространяются по воздуху и в конечном счете попадают в почву, в результате чего подавляют рост других растений. На аэрофотоснимке хорошо видна кайма голой земли вокруг зарослей этого вида. Непосредственно вокруг кустов находится «мертвая зона», а затем идет зона с небольшим числом низкорослых однолетников
Рис. 31-6. Когда млекопитающие (например, мыши) не допускаются в «мертвую зону», однолетники прекрасно растут у самого края кустов, на территории, где они обычно отсутствуют. Эти результаты, представленные Б. Бартоломью из Калифорнийской академии наук, указывают на то, что наличие мертвой зоны может быть полностью обусловлено действием травоядных животных, независимо от того, образуют растения токсины или нет

Взаимоотношения растений и фитофагов
Обширные пространства Австралии были когда-то покрыты колючими зарослями кактуса опунции (Opuntia), занесенного сюда из Латинской Америки. В результате плодородные земли нс могли использоваться для выпаса и хозяйство огромных территорий внутренней части страны сильно страдало. Сейчас опунция почти полностью уничтожена насекомым кактусовой огневкой (Cactoblastis cactorum), открытым в Южной Америке и сознательно завезенным в Австралию. Дело в том, что личинки этого вида питаются кактусами. Кактусовая огневка, некогда обильная в Австралии, сейчас настолько редка, что ее трудно обнаружить, даже специально осматривая немногие оставшиеся заросли опунции, однако нет сомнений, что она продолжает контролировать численность популяции этого растения в Австралии (рис. 31-7).
Рис. 31-7. А. Густые заросли опунции (Opuntia inermis) в смешанном кустарниковом редколесье шт. Квинсленду Австралия, октябрь 1926 г. Б. Тот же самый лес в октябре 1929 г. после того, как кактусы были уничтожены специально завезенной южноамериканской бабочкой огневкой кактусовой. Впервые появившиеся в мае 1925 г. личинки этого насекомого уничтожили кактусы более чем на 120 млн. га пастбищ.

В целом как кратковременное, так и долговременное влияние фитофагов на растения очень велико. Как говорилось в гл. 29, эти взаимоотношения привели в ходе эволюции к возникновению у растений широкого спектра защитных химических веществ, обычно называемых «вторичные растительные метаболиты». Способность растений вырабатывать токсины и накапливать их в своих тканях дает им громадное конкурентное преимущество. В самом деле, такие химические вещества, по-видимому, важнейший фактор, контролирующий численность растительноядных насекомых в природе. Это преимущество аналогично тому, которое дает образование шипов или жестких кожистых листьев, и ученые, занимающиеся проблемой повышения устойчивости культур к фитофагам, ведут интенсивное изучение таких соединений. Учитывая их роль в наземных экосистемах, не приходится удивляться, что недавно вещества со сходными функциями обнаружены и у многих морских водорослей. Показано, что они защищают водоросли от выедания морскими фикофагами.
Взаимоотношения растение — фитофаг и растение — паразит могут быть очень сложными. Например, горох (Pi- sum sativum) хорошо защищен от грибов-паразитов образующимся у него веществом пизатином. Однако многие штаммы важного паразитического аскомицета Fusarium содержат ферменты монооксигеназы, превращающие пизатин в менее токсичное вещество, и в результате могут поражать эту культуру. Люди также используют монооксигеназы для инактивации некоторых потенциально опасных веществ. Таким образом, «химическая война» между растениями и растительноядными животными ведется постоянно.
Защитные вещества, вырабатываемые растениями, часто неприятны на вкус, но некоторые из них отпугивают фитофагов иным способом. Хромены, например, могут влиять на ювенильный гормон насекомых (необходимый для нормального протекания жизненного цикла), т. е. действовать как настоящие инсектициды. Мексиканское растение гелениум (Helenium spp.) вырабатывает геленалин, действующий как сильный репеллент насекомых. Пиретрины, обнаруженные впервые у сложноцветного пиретрума (Pyrethrum), — хороший пример природных инсектицидов, получаемых промышленным путем из близкого к пиретруму рода Chrysanthemum. Даже восковая поверхность листьев, которая трудна для переваривания, может служить хорошим барьером против насекомых и грибов.
Когда растения поражены грибами или бактериями, они часто защищаются, вырабатывая антибиотики, называемые фитоалексинами. Синтез этих липидоподобных веществ может быть стимулирован простым повреждением листьев. По-видимому, они образуются в ответ на присутствие специфических углеводных молекул, называемых элиситорами, входящих в состав оболочек грибных и бактериальных клеток. Высвобождаясь из этих оболочек ферментами пораженного растения, они диффундируют через его клетки, как гормоны, в конечном итоге связываются со специфическими рецепторами плазматических мембран и вызывают метаболические изменения, приводящие к синтезу фитоалексинов. В принципе можно было бы опрыскивать элиситорами непораженные посевы, защищая их таким образом от грибной или бактериальной инфекции. Такой способ был бы аналогичен вакцинации людей.
Однако одна из трудностей состоит в том, что энергетические затраты растения на синтез больших количеств фитоалексинов могут снизить окончательный урожай даже сильнее, чем паразиты. Тем нс менее понимание механизма образования этих веществ растениями очень важно для защиты посевов, и некоторые синтетические элиситоры уже получены и испытываются. Изменение генетической основы устойчивости, ставшее сейчас возможным благодаря методам генной инженерии (см. гл. 30), открывает новые пути повышения сопротивляемости фитофагам, не связанные с увеличением энергетических затрат организма.
Вместо фитоалексинов некоторые растения вырабатывают таннины и другие фенольные соединения, играющие, вероятно, сходную роль в природе. Когда непарный шелкопряд (Lymantria dispar) объедает листья дуба (Quercus spp.), в новых листьях, появляющихся на деревьях, содержание таннинов и других фенольных соединений гораздо выше. Такие листья гораздо жестче и содержат меньше воды, чем те, которые они заменили. Эти различия достаточно велики, чтобы новые условия питания привели к задержке роста личинок и, таким образом, замедлили рост популяции шелкопряда. Таннины, вероятно, затрудняют усвоение пищи насекомыми, взаимодействуя с растительными белками и делая их непереваримыми. Сходные реакции могут наблюдаться и у других растений. Например, когда американский заяц-беляк сильно обгладывает некоторые деревья и кустарники, например, березу бумажную (Betula papyrifera), растения образуют новые побеги, в которых содержание смол и фенольных соединений гораздо выше, чем до повреждения.
Сходные данные были получены Д. Родсом из Вашингтонского университета для ив и ольхи. Более того, Родс обнаружил, что эти деревья, по-видимому, образуют некое летучее вещество при нападении гусениц шелкопряда или экспериментальном удалении значительной доли их листовой поверхности. Оно, вероятно, передается от дерева к дереву по воздуху и действует таким образом, что неповрежденное дерево, не находившееся в непосредственном контакте с поврежденным, реагирует так, как будто подверглось нападению насекомых. В обоих случаях растения вырабатывали повышенное количество фенольных соединений и таннинов, снижающих их съедобность для фитофагов в течение 36 ч с начала поражения (рис. 31-8). Однако это летучее вещество, несмотря на ведущиеся исследования, до сих пор не идентифицировано.
Рис. 31-8. Относительная скорость роста гусениц кольчатого коконопряда, питающихся листьями десяти деревьев ивы Salix sitchensis, экспериментально заселенных колониями этих гусениц 28 мая 1981 г., листьями десяти деревьев того же вида, расположенных поблизости, и листьями двадцати деревьев, растущих вдалеке. Внизу — число гусениц, отмеченных на деревьях. «Аппетитность» ближних деревьев (оцениваемая по их способности поддерживать рост гусениц) с течением времени заметно падала, а у дальних деревьев оставалась неизменной в течение всего эксперимента

То, что растения могут сами защищать себя, вырабатывая токсины, следует учитывать в сельском хозяйстве. Например, дикорастущие виды семейства тыквенных (Cucurbitaceae) образуют в плодах и листьях горькие терпены, отпугивающие большинство фитофагов. У культурных сортов эти вещества уже не вырабатываются, так как селекция шла по пути улучшения вкуса плодов, и растения съедобны для вредителей. Чтобы защитить их, приходится принимать специальные меры, в частности опрыскивать культуры инсектицидами. Разводимые арбузы (Citrullus vulgaris) подвергаются нападению гораздо большего числа насекомых, чем их дикорастущие формы, и, чтобы сохранить урожай, эти фитофаги должны быть нейтрализованы.
Процесс опыления — особая форма взаимоотношений между фитофагами и растением, при которой определенная часть последнего (чаще всего нектар) поедается животным- опылителем. По многим признакам эти отношения являются формой мутуализма, поскольку оба партнера получают от них пользу. В ходе эволюции вырабатывались механизмы привлечения фитофагов, что, как описано в гл. 29, привело к огромному разнообразию цветков покрытосеменных. При такой форме отбора и растения, и животные все лучше адаптировались друг к другу и появлялись все более специализированные системы опыления.
Вторичные растительные метаболиты, поглощаемые фитофагами, в свою очередь могут играть определенную роль в их отношениях с другими животными. Некоторые насекомые, накапливая такие яды в своих тканях, становятся несъедобными для хищников (рис. 31-9). Ряд половых аттрактантов насекомые получают из растений, которыми питаются. Они концентрируют эти вещества, а затем используют их для привлечения партнеров противоположного пола.
Рис. 31-9. А. Бабочка данаида (Danaus plexippus) получает сердечные гликозиды из растений семейства ластовневых (Аsclepiadасеае), на которых живут и питаются ее гусеницы (Б). В результате она несъедобна для птиц и других позвоночных. Она «предупреждает» об этом ярким оранжево-черным узором взрослых особей, а также хорошо различимыми белыми, желтыми и черными полосами на гусеницах. Даже светло- желтые заметные яйца данаиды содержат достаточно сердечных гликозидов, чтобы хищники их не трогали

Муравьи, населяющие шипы акаций, о которых уже шла речь при обсуждении мутуализма, играют в жизни этих растений примерно такую же роль, что и вторичные метаболиты, — они защищают деревья от фитофагов. Интересно, что виды Acacia, обычно нс заселяемые муравьями, содержат горькие вещества, а «виды-муравейники» их не образуют. Получается, что химические соединения и насекомые выполняют одну и ту же функцию.
В целом взаимоотношения организмов в сообществе предельно сложны. Растения, встречающиеся вместе, влияют друг на друга бесконечно разнообразными путями, лишь немногие из которых мы сейчас начинаем понимать. Маловероятно, чтобы для выживания вида была важна судьба отдельной особи; скорее его успех в конкретном местообитании определяется группой связанных между собой особей и типами существующих между ними взаимоотношений.
Круговорот питательных веществ
С точки зрения снабжения питательными веществами (биогенами) экосистема более или менее самостоятельна. Одна из важнейших причин такой автономии — постоянный круговорот химических элементов между организмами и средой. Пути некоторых незаменимых для живого элементов, известные как циклы биогенов, обсуждались в гл. 26. В идеале нс теряется ничего и общий запас питательных веществ постоянно возобновляется и остается доступным для организмов. Скорость потока от неживой природы к организмам и обратно, доступное количество в физической среде и форма присутствия в этой среде у разных элементов неодинаковы.
круговорот в лесной экосистеме.
Изучение экосистемы листопадных лесов показало, что растения в этом сообществе являются основными накопителями биогенов. Исследования были проведены в экспериментальном лесу Хаббард-Брук (национальный парк «Уайт-Маунтин», шт. Нью-Гэмпшир). Сначала был разработан способ определения «минерального бюджета», т. с. поступления и потерь элемента, в различных участках сообщества. Анализируя содержание биогенов в дожде и снеге, можно оценить первый параметр, а построив бетонные плотины, собирающие воду с выбранных делянок, — второй (рис. 31-10). Место было очень удобно для проведения такого эксперимента: подстилающая скальная порода находилась сразу же под поверхностью почвы, так что из нее вымывалось очень мало веществ (почвенные воды просачивались на небольшую глубину).
Рис. 31-10. Плотина в экспериментальном лесу Хаббард-Брук (шт. Нью-Гэмпшир). Вода от каждой из шести изучаемых экосистем собиралась в запрудах (таких, как показанная на снимке), когда стекала с территории, и ее химический состав анализировался. Деревья и кустарники на площади выше этой плотины были срублены. Эксперименты показали, что такое сведение леса значительно увеличивает потери питательных веществ экосистемой

Исследователи установили, что коренной лес чрезвычайно эффективен с точки зрения сохранения минеральных элементов. Так, ежегодные чистые потери кальция из этой экосистемы составили 9,2 кг/га, т. е. всего 0,3%. Азот экосистема накапливала в количестве 2 кг/га в год. Близким, хотя и несколько меньшим было чистое поступление калия.
В лесу Хаббард-Брук биологическая регуляция круговорота биогенов была проверена следующим образом. Зимой 1965/66 г. все деревья, подрост и кустарники в водосборном бассейне площадью 15,6 га были вырублены. При этом органический материал не удалялся, и почва не нарушалась. В течение следующей весны территория была опрыскана гербицидами, чтобы подавить рост растений. В течение четырех месяцев, с июня по сентябрь 1966 г., поверхностный сток с этой площади был в четыре раза выше, чем в предыдущие годы. Чистые потери кальция в 20, а калия в 21 раз превысили наблюдаемые в ненарушенном лесу. Наиболее серьезные изменения наблюдались в цикле азота. Ткани мертвых растений и животных продолжали разлагаться до аммиака (или ионов аммония), которые превращались нитрифицирующими бактериями в нитраты, т. е. в форму, в которой этот элемент обычно ассимилируется растениями. Однако в отсутствие последних нитраты не удерживались в почве. Чистые потери азота в среднем достигли в 1966 — 1968 гт. 120 кг/га в год. Побочным результатом стало водорослевое «цветение» дренирующих территорию водотоков, загрязненных нитратами, концентрация которых здесь превысила безопасный уровень для питьевой воды, установленный Службой здравоохранения США.
Азот не всегда теряется из нарушенных лесных экосистем так быстро, как в лесу Хаббард-Брук. Реальная скорость этого процесса зависит от особенностей его цикла в конкретном сообществе до нарушения. Например, если потребности микроорганизмов в азоте, высвобождаемом после вырубки леса, велики, то потери могут и не быть такими большими, как в рассмотренном нами случае.
ТРОФИЧЕСКИЕ УРОВНИ
Кроме физических (неживых) компонентов каждая экосистема включает два биотических (живых) — автотрофов и гетеротрофов. Автотрофы — главным образом фотосинтезирующие организмы, способные использовать солнечную энергию для создания собственных органических веществ. Поскольку гетеротрофы этого не могут, они должны использовать органические молекулы, образованные автотрофами. Гетеротрофы составляют несколько пищевых, или трофических, уровней. Это, во-первых, первичные консументы (растительноядные животные), непосредственно питающиеся автотрофами. Во-вторых, это вторичные консументы (плотоядные и паразиты), потребляющие других животных. Наконец, деструкторы (редуценты), включая грибы, бактерии и различных мелких животных, разлагают органические вещества до неорганики. Все эти уровни присутствуют в большинстве экосистем.
Организмы всех трофических уровней образуют то, что называется пищевой цепью (рис. 31-11). Отношения между организмами в пищевых цепях регулируют поток энергии в экосистеме. Длина и сложность таких цепей сильно варьируют. Обычно каждый организм имеет несколько источников питания и сам употребляется в пищу несколькими другими организмами. В большинстве случаев правильнее говорить о пищевой сети (рис. 31-12). Сложность трофических взаимоотношений оказывает существенное влияние на свойства экосистемы в целом.
Рис. 31-11. Пищевая цепь. Каролинская коробчатая черепаха (Terrapene Carolina triunguis) ест улитку, питающуюся грибами, которые в свою очередь разлагают органическое вещество почвы
Рис. 31 -12. Пищевые связи сельди (Сluреа harengus) в Северной Атлантике, иллюстрирующие сложность пищевой сети

Поток энергии
В экосистеме этот поток обычно начинается с солнечной радиации, улавливаемой в процессе фотосинтеза и используемой для образования молекул углеводов. Энергия не циркулирует в экосистемах, а передается от автотрофов (как правило, фотосинтезирующих организмов, т. е. растений, водорослей и некоторых бактерий) к консументам (животным и гетеротрофным протистам), а затем к деструкторам. На каждом этапе основная ее часть рассеивается в форме тепла, в конечном итоге возвращающегося в мировое пространство в виде инфракрасного излучения.
На планете ежегодно производится огромное количество биомассы. Этим термином обозначают органическое вещество в целом, включая одревесневшие части деревьев, запасные питательные вещества, кости и т. д. В настоящее время глобальная продукция биомассы оценивается в 200 млрд, т/год. Несмотря на эту огромную цифру, эффективность фотосинтезирующих организмов с точки зрения превращения солнечной энергии в органическое вещество невелика. Растением обычно используется менее 1% падающего на него светового излучения (см. гл. 5). Однако некоторые особенно продуктивные растительные сообщества и водные системы могут ежегодно превращать в химическую энергию до 3% воздействующей на них солнечной радиации.
Когда органическое вещество, синтезированное растениями, поглощается фитофагами, энергия высвобождается. В основном она теряется в виде тепла, но часть потребленной органики превращается в животную ткань. Как правило, только 2 — 10% усвояемой энергии растения добавляется к массе фитофагов — остальное теряется на дыхание. Сходные отношения потребления и потерь обнаруживаются и на каждом последующем трофическом уровне. Так, если ежедневно растения воспринимают в среднем 1500 кал солнечной энергии на 1 м2 поверхности суши, то в растительное вещество превращается всего около 15 кал. Из этого количества около 1,5 кал образует тело фитофагов и примерно 0,15 кал — тело плотоядных, питающихся фитофагами.
Л. Кол из Корнеллского университета, исследуя озеро Каюга, подсчитал, что из каждых 1000 кал солнечной энергии, использованной водорослями, около 150 кал переходит в мелких водных животных и 30 кал — в корюшку (мелкая рыбка). Если съесть эту корюшку, можно получить 6 кал из первоначальных 1000, усвоенных водорослями. Однако если форель съест корюшку, а мы затем съедим форель, то нам достанется всего 1,2 кал из этих 1000. Очевидно, что мы получили бы больше энергии, если бы съели корюшку, а нс форель, но форель — деликатес, а корюшка — менее привлекательная для человека пища. Во время голода людям следует полностью переходить на растительную пищу, не допуская тем самым десятикратных потерь энергии за счет скармливания растений животным. Чтобы максимально использовать солнечную радиацию, улавливаемую растениями, мы должны стать фитофагами.
Пищевые цепи обычно ограничены тремя или четырьмя звеньями; количество пищи, остающееся в конце более длинной цепи, настолько мало, что может обеспечить очень мало организмов. Размер тела также влияет на структуру пищевых цепей. Например, животные определенного звена обычно должны быть достаточно крупными, чтобы нападать на животных предыдущего трофического уровня. Однако большая часть биомассы в экосистеме используется деструкторами, в частности грибами и бактериями, обеспечивающими круговорот биогенов.
Как следует из отношений, которые только что обсуждались, общая биомасса с переходом к следующему трофическому уровню экосистемы обычно резко уменьшается, и возникает так называемая «пирамида массы» (рис. 31-13). Эти отношения иногда не выдерживаются, если существует быстрое обновление популяций первичных продуцентов, например, водорослей в озере. Скорость обновления приобретает в таких случаях решающее значение, и общая биомасса этого уровня в каждый отдельный момент времени может быть относительно низкой. Если измерить энергию, то оказывается, что она так же быстро уменьшается с увеличением трофического уровня, как и биомасса, т. е. существует «пирамида энергии» с гораздо меньшей се суммой в телах всех хищников, присутствующих в данном сообществе, чем, например, во всех растениях. Как правило, на нижних трофических уровнях всегда намного больше особей, чем на высоких, т. е. можно говорить и о «пирамиде чисел». Из этого также следует, что если все организмы в экосистеме разделить на классы размеров, то мелкие животные будут гораздо многочисленнее крупных.
Рис. 31-13. Пирамиды массы (А), энергии (Б) и чисел в различных сообществах. На каждый более высокий уровень переносится относительно небольшое количество массы и энергии

Практический аспект изучения потоков энергии в экосистемах связан с усилиями человечества создать из растений возобновляемые источники энергии (так называемое биологическое превращение энергии). Подсчитано, что огромное количество отходов, получаемое ежегодно после сбора урожая или вырубок леса, может дать энергию, эквивалентную 1% бензина или 4% электроэнергии, потребляемых в США за год. Однако этот потенциал ограничен энергетическими затратами на сбор материала. Посадка быстро растущих деревьев и других растений могла бы обеспечить основной возобновляемый источник энергии в будущем и стать одним из самых эффективных способов использования солнечной энергии.
Развитие сообществ и экосистем
Сукцессия
Некоторые растительные сообщества остаются стабильными многие годы, в то время как другие быстро изменяются. В последнем случае в их составе происходят закономерные смены, которые называются сукцессией. Вырубка в лесу быстро заселяется окружающими деревьями; сходным образом луг в конечном итоге сменяется лесом. Аналогичные явления наблюдаются и в природных открытых местообитаниях — в озерах, на скальных склонах и т. д. Процесс сукцессии непрерывен и идет по всей планете.
Скорость сукцессии на всех временно незанятых территориях различна. Некоторые озера, например, постепенно заполняются остатками водных растений, выступающие из воды виды формируют почву, и водоем превращается в луг, на котором может появиться влаголюбивый кустарник и, наконец, развиться характерный для этой природной зоны лес (рис. 31-14). Скалы под действием замерзания и оттаивания и других физических факторов выветриваются и разрушаются; этот процесс иногда ускоряется лишайниками, выделяющими определенные химические вещества, непосредственно разъедающие породу. Вокруг оснований лишайников накапливается почва, после чего они уступают место мхам и цветковым растениям (рис. 31-15). Корни цветковых растений раздвигают трещины в скале, разрушая ее все сильнее. В конечном итоге, возможно через сотни лет, этот процесс измельчения дает почву, заселяемую лесом или другой характерной для данного района растительностью. На ранних стадиях сукцессии часто преобладают растения, образующие симбиоз с азотфиксаторами. Другие примеры показаны на рис. 31-16.
Рис. 31-14. А. Полупогруженная растительность вдоль края озера. Б. Растения с плавающими листьями, например, кувшинка (Nymphaea odorata), распространяясь по всей поверхности водоема, постепенно заглушают донные виды. В. Водяной гиацинт (Eichhornia odorata) играет аналогичную роль в более теплых областях. Г. Болотные злаки, осоки и рогозы (Тypha sрр.) на месте заросшего озера завершают сукцессию

Рис. 31-15. Ранняя стадия сукцессии на скалистом склоне. Лишайники начинают разрушать породу, а папоротники и бриофиты накапливают почву в небольших трещинах
Рис. 31-16. А. Молодые деревья пихты бальзамической (Abies balsamea), растущие под тополем (Populus tremuloides) и вытесняющие его, в северной Миннесоте; стадия лесной сукцессии, ведущей к климаксовому сообществу ели белой (Picea glauca) и пихты бальзамической. Б. Всход клена красного (Acer rulyrum), поднимающийся над хвоей веймутовой сосны (Pinus strobus). Спелые сосны этого вида пропускают так мало света, что их собственные всходы под ними не выживают и сохраняется только подрост таких теневыносливых видов, как клены и дубы, способных там закрепиться. С другой стороны, сукцессия тормозится из-за подавления почвенных азотфиксирующих бактерий химическими веществами, вымываемыми из опавшей сосновой хвои

Любая экосистема проходит различные стадии сукцессии до тех пор, пока нс возникает «зрелое», так называемое климаксовое сообщество. Хотя характер этого конечного состояния зависит от климата, климаксовые сообщества часто гораздо стабильнее тех, на смену которым они появились. Организмы в них образуют очень сложную систему взаимоотношений.
Некоторые аспекты сукцессии имеют огромное значение для людей. Например, когда европейские поселенцы впервые прибыли в большом количестве в Калифорнию, они увидели там вдоль большей части массива Сьерра-Невада величественные леса из сосны Ламберта (Pinus lambertiana). Позже, несмотря на то что сторонники охраны природы пытались превратить некоторые из этих лесов в национальные парки, многие сосновые сообщества были замещены другими породами, в частности пихтой одноцветной (Abies concolor) и кедром ладанным (Calocedrus deccurrens). Почему же это произошло?
Дело в том, что сосна Ламберта была на данной территории элементом определенной стадии лесной сукцессии, причем эта стадия поддерживалась периодическими пожарами. После появления европейцев масштабы и частота пожаров значительно уменьшились. Без периодических слабых прореживающих пожаров, «пробегавших» по соснякам, возникли такие густые заросли кустов и невысоких деревьев, что светолюбивые сосны не смогли возобновляться. Только система контролируемых палов способна сохранить оставшиеся леса из сосны Ламберта в их исходном состоянии (рис. 31-17, Б).
Рис. 31-17. А. После лесного пожара начинается вторичная сукцессия, т. е. восстановление растительности от ближайших неповрежденных огнем участков. Некоторые растения дают поросль от своих оснований, другие рас
сеивают на гарь большое количество семян. В одной группе сосен шишки не раскрываются и не рассыпают семена до тех пор, пока на них не подействует огонь (см. также рис. 19-7). Б. Сосна Ламберта в южной части массива Сьерра-Невада в Калифорнии. После прекращения (благодаря усилиям людей) лесных пожаров, этот вид замещается другими деревьями, например, кедром ладанным (Calocedrus decurrens) (огромное дерево справа на снимке)

Реколонизация
Когда люди преобразуют ландшафт, происходят изменения в структуре сообществ. На заброшенных полях, голых песчаных дюнах, на улицах покинутых поселков американского Запада происходит сукцессия в направлении экосистем, более или менее напоминающих существующие на соседних, менее нарушенных территориях (рис. 31-18). Если времени достаточно, сукцессионный процесс может постепенно привести к восстановлению коренной растительности. Например, считается, что в северных лиственных лесах Северной Америки и Евразии для восстановления растительной биомассы и питательных веществ, удаленных из них в результате вырубки деревьев, требуется от 60 до 80 лет. В других сообществах этот процесс может идти быстрее или медленнее. В любом случае для его успеха требуются значительное время и источник семян для реколонизации. Во многих регионах, особенно в тропиках, этих условий нет из-за сильнейшего воздействия, оказываемого на растительность быстро растущим населением, и общей бедности (см. гл. 30).
Рис. 31-18. А. Лес с сосной ладанной (Pinus lueda) на экспериментальной делянке научной станции «Толл-Тимберс» около Таллахасси (Флорида). Последний контролируемый пал на этой территории был проведен 23 марта 1967 г. Б. В отсутствие пожаров здесь быстро идет сукцессия. Фотография сделана на том же месте в 1982 г.

После природных катастроф реколонизация приводит к таким же изменениям. Например, в августе 1883 г. сильное вулканическое извержение уничтожило половину острова Кракатау в Зондском проливе (40 км от Явы), а оставшаяся его половина покрылась слоем пемзы и пепла толщиной более 31 м. Были засыпаны и соседние острова, так что вся растительность и животный мир на них погибли. Однако вскоре после этого началась реколонизация Кракатау, и ожидавшееся число (исходя из первоначального разнообразия орнитофауны) — около 30 видов сухопутных и пресноводных птиц — восстановилось здесь в течение примерно 30 лет. Быстро происходило и заселение растениями: уже к 1934 г. на острове было зарегистрировано более 270 их видов.
В шт. Вашингтон (США) при сильном извержении вулкана Сент-Хеленс 18 мая 1980 г. мощная лавина вулканического материала с вершины и северной стороны горы сошла в долину реки Норт-Таутл. В течение 15 мин более 61 000 га леса и рекреационных территорий были опустошены боковой ударной волной, вывалившей древостой на 21 000 га и погубившей деревья и другие растения, оставив их на корню, еще на 9700 га. Кроме того, девятичасовое извержение покрыло всю площадь полуметровым слоем пепла, пемзы и камней, разбросанных этой волной (рис. 31-19).
Рис. 31-19. Последнее извержение вулкана Сент-Хеленс в шт. Вашингтон произошло 18мая 1980 г. А. Продуктивные леса из ду гласии (Pseudotsuga menziesii), тсуги западной (Tsuga heterophylla) и пихт Abies amabilis и А. procera, вываленные боковой ударной волной. Б. На площади более 60 км1 отложился мощный слой вулканического материала. В. Летом 1980 г. из-под пепла проросло большое количество многолетних трав, в частности иван-чай (Epilobium angustifolium). Эта фотография сделана через год после извержения. Г. Дугласия и многие другие растения с семенами, распространяемыми ветром, реколонизировали участки, покрытые таким глубоким слоем вулканических отложений, что растения, погребенные под ними, погибли. Эта фотография сделана в 1984 г.
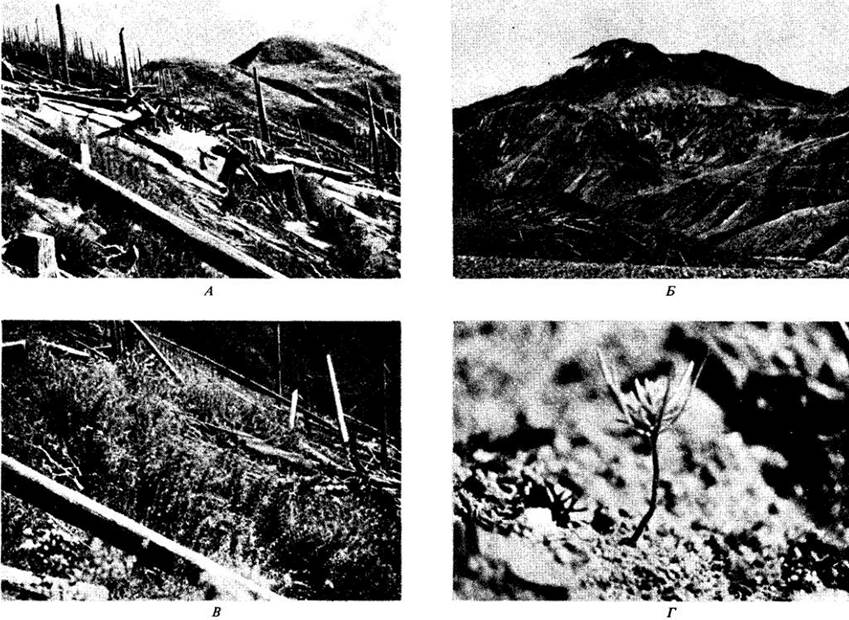
Жизнь стала возобновляться на склонах, пострадавших от извержения, почти сразу, но растения смогли пробиться сквозь толщу вулканических отложений только следующей весной. Большая часть этих отложений была вскоре размыта, и сюда попали распространяемые ветром семена и плоды. Такой занос был особенно важен для территорий, покрытых лавинным материалом и застывшей вулканической лавой с пеплом настолько сильно, что растения, погребенные под ними, погибли. Выжили многие мелкие животные как под землей, так и в озерах и ручьях; позвоночные вскоре также вернулись на эти земли. Периоды между извержениями этого вулкана составляли за последние 35 веков 100 — 150 лет, поэтому сам характер повреждений и восстановительные процессы, наблюдавшиеся учеными после 1980 г., типичны для природных явлений, периодически влияющих на живые системы в вулканических районах.
События, последовавшие за извержением Сент-Хеленс, — яркий пример сукцессии, однако не следует забывать, что она типична для всех сообществ и происходит во всем мире. Это один из факторов, поддерживающих разнообразие живого на нашей планете.
Заключение
Экосистема — высший уровень биологической сложности и единства. Это приводимый в действие энергией солнца самоподдерживающийся комплекс, в котором происходит регулируемый круговорот необходимых биогенов. В пределах экосистем сообщества организмов взаимодействуют с физической средой.
Некоторые из взаимосвязей существ внутри сообществ могут быть объединены в три основные группы: мутуализм, конкуренция и отношения растение — фитофаг. Мутуализм полезен обеим взаимодействующим популяциям. Примеры его — образование лишайников, развитие азотфиксирующих бактерий в клубеньках на корнях бобовых, микоризные ассоциации грибов и корней сосудистых растений, тесная взаимоприспособленность некоторых цветковых и их опылителей.
Конкурентные отношения установлены для большинства обитающих вместе растений. Одной из важнейших форм является конкуренция за свет. Растения также вырабатывают защитные химические вещества, подавляющие рост по соседству других видов. Такие аллелопатические отношения играют большую роль в определении будущей структуры сообществ.
Фитофаги регулируют размножение растений, уничтожая их фотосинтезирующие, запасные или репродуктивные органы. Растения противодействуют этим повреждениям, приобретая в процессе эволюции шипы, жесткие листья или, что наиболее важно, химические средства защиты. Когда насекомое вырабатывает устойчивость против растительных токсинов, оно получает не только новый и часто почти нетронутый источник пищи, но может также использовать эти производимые растением вещества для защиты от собственных врагов. Взаимосвязи в процессе опыления — особый случай отношений растение — фитофаг, при котором привлечение последнего важнее его отпугивания. Некоторые отношения растений с фитофагами в значительной степени мутуалистичны, например, облигатные связи между шиповатыми акациями и обитающими на них муравьями на востоке Мексики.
Экосистема состоит из неживых элементов и двух типов живых существ — автотрофов и гетеротрофов. Среди гетеротрофов различаются первичные консументы (растительноядные), вторичные консументы (плотоядные и паразиты) и, наконец, деструкторы. Организмы этих трофических уровней объединены в пищевые цепи или сети.
Проходя через экосистемы, примерно 0,1 — 1% падающей солнечной энергии превращается зелеными растениями в энергию химических связей. Когда растения поедаются животными, около 2 — 10% их энергии накапливается на следующем трофическом уровне; со сходной эффективностью идет и дальнейшая передача энергии по пищевой цепи. Количество ее, остающееся после нескольких таких переходов, настолько мало, что пищевые цепи редко включают более трех или четырех звеньев.
Сукцессии происходят на опустошаемых искусственным или естественным путем территориях или при появлении новых субстратов, например, застывшей вулканической лавы, скальных обнажений, песчаных или гравийных островов на реке. В ходе сукцессии виды растений и животных постепенно сменяют друг друга; некоторые из них характерны только для ранних ее стадий, другие — только для более поздних. Количество органического вещества, накопленного организмами в процессе сукцессии, возрастает, разнообразие видов в экосистеме значительно увеличивается, а отношения между ними становятся все более сложными. Пик продукции наблюдается довольно рано, а на более поздних стадиях быстро усиливается процесс дыхания. В конечном итоге сукцессия приводит к климаксовому сообществу, воспроизводящему себя неопределенно долго, если не происходит каких-то глубоких изменений среды.
Приложение 1. Пестициды и экосистемы
Только в США для обработки посевов ежегодно производится приблизительно 0,5 млрд. т пестицидов. Подсчитано, что из этого огромного количества только около 1% фактически воздействует на вредные организмы, а остальное либо проникает в почву, воду и другие организмы той же экосистемы, либо переносится в соседние экосистемы, что может крайне отрицательно сказываться на их функционировании. Например, не исключена гибель определенных видов, что иногда ведет к нарушению функционирования экосистемы в целом или косвенно к исчезновению других видов. Популяции важнейших деструкторов, например, дождевых червей или других почвенных организмов, могут под действием пестицидов настолько сократиться, что экосистема как таковая прекратит свое существование. Интенсивность такого воздействия зависит от токсичности химических веществ и от их устойчивости в окружающей среде.
Одна из проблем, связанных с некоторыми химическими загрязнителями, состоит в том, что они имеют тенденцию концентрироваться по мере прохождения по пищевым цепям, достигая максимального содержания в их конечных звеньях — в теле хищных животных. Например, хлорорганические пестициды, в частности ДДТ (в США и других развитых странах он в настоящее время запрещен законом), накапливаясь в тканях хищных птиц, служили причиной ненормального истончения скорлупы откладываемых ими яиц. Такие яйца часто раскалывались задолго до вылупления птенцов, что заметно сокращало плодовитость видов. Кроме того, многие популяции насекомых, бактерий и грибов выработали устойчивость к пестицидам, предназначенным для борьбы с ними. По приблизительной оценке, из 2000 видов основных насекомых-вредителей около четверти уже включают популяции, нечувствительные к одному или нескольким инсектицидам. Точно так же у ряда сорных растений появилась устойчивость к гербицидам (см. гл. 30).
Другие воздействия нс столь прямолинейны. Например, полезные хищники, поддерживающие в естественных условиях низкий уровень популяций вредителей, могут погибнуть в результате отравления пестицидами, что приведет к вспышке численности тех видов, против которых применялись эти химикаты. Как и в большинстве случаев антропогенного вмешательства, общий результат применения пестицидов заключается в снижении разнообразия экосистем, которые ими затронуты. Несмотря на эти осложнения, высокоурожайное сельское хозяйство сейчас существенно зависит от применения таких химикатов.
Однако в связи с тем, что их воздействие зачастую приводит к драматическим последствиям, ученые активно ищут альтернативные способы защиты растений. К ним относятся селекция культур, устойчивых к вредителям (при этом будут полезны методы генной инженерии, описанные в предыдущей главе), разработка менее токсичных и менее стойких пестицидов, чем используемые в настоящее время, комплексные системы борьбы с вредителями, включающие использование хищников и болезней вредителей, а также рациональные пестицидные обработки.
Приложение 2. Системы защиты растений семейства Пасленовых
Многие растения из семейства пасленовых (Solanaceae), к которому относятся такие экономически важные для человека культуры, как картофель (Solarium), томаты (Lycopersi- соп) и табак (Nicotiana), в качестве одного из основных средств защиты от вредных насекомых выработали железистые волоски, выделяющие липкие, а в некоторых случаях и токсичные жидкости. У разводимого картофеля (Solarium tuberosum) таких волосков нет, но некоторые его дикие родственники (например. Solarium berthaultii) густо покрыты ими на листьях и стебле. Приклеиваясь к липким выделениям, насекомые как бы попадают в ловушку и постепенно умирают от истощения. Этот защитный механизм эффективен против многих их видов, включая таких важнейших вредителей картофеля, как тли, земляные блошки и цикадки. В настоящее время в нескольких странах предпринимаются попытки получить такие волоски и у высокоурожайных сортов культурного картофеля, который легко скрещивается с S. berthaultii.
Железистые волоски — только один из элементов защитного арсенала пасленовых. Листья представителей этого семейства богаты такими алкалоидами, как никотин, атропин и гиосциамин, которые ядовиты для многих фитофагов и обусловливают галлюциногенные свойства дурмана обыкновенного (Datura stramonium) и других пасленовых. Стероидные гликозиды, найденные в их листьях, также предохраняют растения от поедания многими фитофагами, но одновременно стимулируют к нему многих насекомых, специализированных на питании растениями этого семейства. Как уже упоминалось в предыдущей главе, некоторые виды Solanum изучаются в настоящее время как возможный источник стероидных молекул для использования в производстве противозачаточных средств. Кроме того, как показано К. Нелсон из Университета шт. Вашингтон, томаты и картофель при сильном одиночном повреждении вырабатывают в течение четырех часов два различных высококонцентрированных ингибитора протеиназ, два фермента, нарушающие пищеварение у фитофагов. Концентрации ингибиторов остаются без изменений еще 5 ч, после чего резко падают.
На фотографии, сделанной с помощью сканирующего электронного микроскопа (А), показан ловчий волосок на листе Solanum berthaultii. Липкое вещество выделяется, когда его четырехстворчатая головка вскрывается при соприкосновении с насекомым. Другие снимки, полученные с использованием такого же микроскопа (Б, В, Г), показывают (с возрастающим увеличением) тлю Myzus persicae, приклеенную к стеблю 5. berthaultii липким веществом из этого волоска.
