МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 5. ВЕГЕТАТИВНЫЕ ОРГАНЫ РАСТЕНИЙ
5.5. Общие закономерности строения вегетативных органов
При всем разнообразии строения вегетативных органов для всех их характерен ряд общих закономерностей. К ним относятся наличие определенного типа симметрии, свойства полярности, способность к метаморфизированию, редукции и абортированию, регенерации.
Симметрия (от греч. symmetria — соразмерность) — широко распространенное в природе явление, характерное как для отдельных органов, так и для целых организмов. В морфологии растений под симметрией понимают возможность разделить орган на несколько зеркально подобных половинок. Плоскость, которая делит орган на симметричные части, называется плоскостью, или осью, симметрии. Вегетативные органы могут быть моносимметричными, бисимметричными, полисимметричными (радиально-симметричными) и асимметричными.
Через моносимметричный орган можно провести только одну плоскость симметрии, следовательно, орган можно разделить только на две зеркально подобные половинки. Моносимметричными являются листья целого ряда растений (сирень обыкновенная — Syringa vulgaris, береза повислая — Betula pendula, копытень европейский — Asarum europaeum). Изредка встречаются моносимметричные стебли (крылатый стебель чины лесной — Lathyrus sylvestris) и корни (досковидные корни фикусов).
Бисимметричными являются уплощенные стебли, через них можно провести две плоскости симметрии (мятлик сплюснутый — Poa compressa, опунция многоколючковая — Opuntia polyacantha).
Если через орган можно провести более двух плоскостей симметрии, он является полисимметричным. Полисимметричны круглые стебли (подсолнечник однолетний — Helianthus annuus), корни (тыква обыкновенная — Cucurbita pepo), корнеплоды (редька посевная — Raphanus sativus, свекла обыкновенная — Beta vulgaris), корневые шишки некоторых растений (чистяк весенний — Ficaria verna, аспарагус густоцветковый «Шпренгера» — Asparagus densiflorus «Schprengeri»), унифациальные листья (очиток едкий — Sedum acre, лук репчатый — Allium cepa), столоны (картофель — Solanum tuberosum).
Особым типом строения органов растений является асимметрия. Через асимметричные органы нельзя провести ни одной плоскости симметрии. Асимметричны листья вязов (вяз гладкий — Ulmus laevis, вяз шершавый — Ulmus scabra), некоторых бегоний (бегония королевская — Begonia rex).
Полярность (от лат. polus, греч. polos — полюс) — одна из общих закономерностей, присущих не только всему растительному организму, но и отдельным его органам, а также клеткам. Полярность характеризуется наличием морфологических и физиологических различий на противоположных концах тела растения или его элементов.
Первые представления о полярности были заложены работами немецкого физиолога растений Г. Фехтинга (1847—1918), который в 1878—1882 гг. провел ряд опытов и обнаружил, что у черенка корни всегда формируются только на базальном конце. Последующие опыты, проведенные с черенками ивы, подтвердили его данные. Как бы не ориентировали черенки, корни всегда образовывались только на морфологически нижнем конце побега, а боковые побеги всегда отрастали на морфологически верхнем (рис. 130).
Рис. 130. Проявление полярности у черенков ивы (Salix sp.): А—нормально ориентированный черенок; Б—черенок в перевернутом состоянии: 1—базальный полюс (основание черенка); 2—апикальный полюс (верхняя часть черенка). Корни образуются только на базальном полюсе
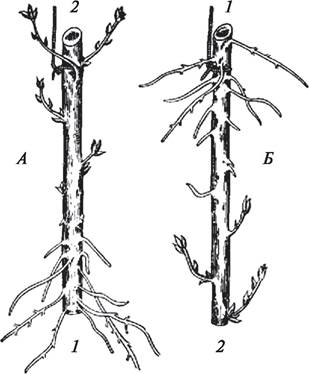
Свойство полярности у растений проявляется уже на ранних стадиях развития, фактически с момента деления зиготы. При первом делении зиготы образуются две клетки — базальная и апикальная, из которых в дальнейшем формируются различные части зародыша будущего нового организма. При дифференцировке зародыша на диаметрально противоположных полюсах образуются конусы нарастания стебля и корня. Присуща полярность корням и листьям, у них имеются четкие различия верхушек и оснований. Благодаря свойству полярности органы растений определенным образом ориентированы в пространстве. Процесс поляризации очень сложен и не до конца изучен. Несомненно, полярность обусловлена физиологическими процессами, которые протекают в растении, и связана в первую очередь с действием
фитогормонов. У высших растений полярность строго детерминирована и сохраняется неизменной даже при изменении условий существования.
Однако у более примитивных организмов (водорослей) в условиях эксперимента было установлено, что при изменении экологических факторов (свет, тепло, влажность) или при механических воздействиях (центрифугирование) могут меняться гормональные и трофические процессы, что влечет за собой изменение поляризации.
Все вегетативные органы способны к метаморфозам, о чем было сказано в соответствующих разделах. Наибольшее разнообразие метаморфизированных структур характерно как для побегов в целом, так и для их компонентов — листьев. Корни, находящиеся в относительно стабильных условиях существования, метаморфизируются реже, причем метаморфозы корней у автотрофных наземных растений связаны главным образом с выполнением запасающей функции.
В процессе морфологической эволюции происходило не только морфофизиологическое усложнение различных органов, но под влиянием условий существования у некоторых видов произошла редукция или даже абортиро- вание отдельных органов или их частей.
Под редукцией (от лат. reductio — возвращение, отодвигание назад) следует понимать уменьшение размеров или числа органов, упрощение их строения и часто связанное с ним изменение присущих им функций. У корневищ листья редуцировались и превратились в сухие чешуи (пырей ползучий — Elytrigia repens, осока мохнатая — Carex hirta). У луковиц лука репчатого сочные чешуи — это основания листьев. Редуцированы листья у ряда сапротрофных растений (подъельник обыкновенный — Hypopitis monotropa, гнездовка обыкновенная — Neottia nidusavis, петров крест чешуйчатый — Lathaea squamaria), а также у некоторых растений-ксерофитов (дифазиаструм сплюснутый — Diphasiastrumcomplanatum, туя западная — Thuja occidentals, кипарис вечнозеленый — Cupressus sempervirens).
У многих водных растений редуцированы корни, у них отсутствуют корневые волоски и корневой чехлик. Редуцированы корни также у растений- паразитов (повилика европейская — Cuscuta europaea) и полупаразитов (омела белая — Viscum album). У раффлезии Арнольди (Rafflesia arnoldii), которая распространена во влажных тропических лесах Индонезии и Филиппин и паразитирует на корнях тетрастигмы (Tetrastigma) и циссуса (Cissus) — родов из семейства Виноградовые (Vitaceae), редуцирован корень и абортированы стебель и листья.
При абортировании (от лат. abortus — выкидыш) орган полностью исчезает. Так, у папоротника сальвинии плавающей (Salvinia natans) — абортирован корень. У повилик (Cuscuta)абортированы листья. Редукция и абортирование органов, как и метаморфозы, — адаптивные процессы, ответная реакция растения на условия существования. Часто термины «редукция» и «абортирование» в ботанической литературе употребляют как синонимы.
Общим свойством вегетативных органов растений является и способность к регенерации (от позднелат. regeneratio — возрождение, возобновление), т. е. к восстановлению утраченных частей организма. Регенерация лежит в основе вегетативного размножения растений. Она может происходить в естественных условиях, но может быть получена в условиях эксперимента. Способность к регенерации у разных таксонов различна. Чем выше степень морфолого-анатомической дифференциации растения и его органов, тем слабее у них способность к регенерации. Регенерация происходит благодаря восстановлению меристематической активности клеток паренхимы и их последующей дифференциации в ткани вегетативных органов.
Способность к регенерации имеет большое практическое значение и широко используется в садоводстве, цветоводстве, биотехнологии.
Аналогичные общие закономерности строения присущи в некоторой степени и репродуктивным структурам.