БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
4. МОРФОЛОГИЯ И АНАТОМИЯ ЛИСТОСТЕБЕЛЬНЫХ РАСТЕНИЙ
В предыдущих главах клетки были рассмотрены как элементарные единицы жизни и конструктивные элементы тканей. В многоклеточном организме каждая клетка также является элементарной единицей живого, но не отдельным организмом. Морфологическое строение многоклеточного организма столь же отличается от его клеточной структуры, как архитектура здания — от устройства кирпичей и других строительных деталей. Можно заниматься морфологией, вообще не имея представления о клетках (что долго и делали). Сложное внешнее строение организма можно представить себе и без дифференциации его тела на отдельные клетки (см. бокс 4.1; рис. 4.1; 11.73, Б; 11.91, а также раздел 2.2.3.6). Однако редкость крупных одноклеточных существ и огромное разнообразие многоклеточных организмов ясно показывают, что многоклеточность представляет собой лучшую основу для эволюции более крупных организмов, чем увеличение и усложнение структуры одной клетки. Формирование многоклеточных организмов предполагает не только увеличение массы и агрегацию, но и упорядоченную дифференциацию первоначально одинаковых клеток Дифференциация и функциональная специализация генетически одинаковых клеток организма (соматических клеток, от греч. soma — тело) основывается на дифференциальном активировании генов (см 2.2.3; 7.2.2.3); каждой возникающей клетке передаются сигналы с информацией о направлении ее последующей дифференциации. Многоклеточный организм возникает как гармоничный ансамбль всех составляющих его клеток и межклеточных сигналов (см. 7.4). Тем самым в качестве биологической единицы выступает уже не отдельная клетка, а надклеточный функциональный «союз» многоклеточного вегетативного тела, так называемая бластема (от греч. blastema — структура; росток, побег). Именно системная целостность отличает многоклеточный организм, или бластему, от простого объединения клеток (ценобия, см. раздел 5 1).
Соматические клетки являются элементами этого целого и выполняют ограниченное число функций. Они могут проявлять себя как элементарные организмы лишь после изолирования из бластемы. Нарушения межклеточных коммуникаций в многоклеточной системе приводят к ненормальным проявлениям роста и дифференциации, например, к образованию опухоли
4.1. Морфология и анатомия
Сетчатка каждого глаза человека связана с головным мозгом примерно миллионом нервных клеток, а передающая способность оптического нерва у человека достигает 100 млн бит в секунду. Однако от простого созерцания к восприятию и умению различать ведет не только прекрасно развитый орган зрения, но и способность нервных центров обрабатывать информацию. При этом понимание структуры объектов есть исходный пункт любого исследования природы, в особенности биологического.
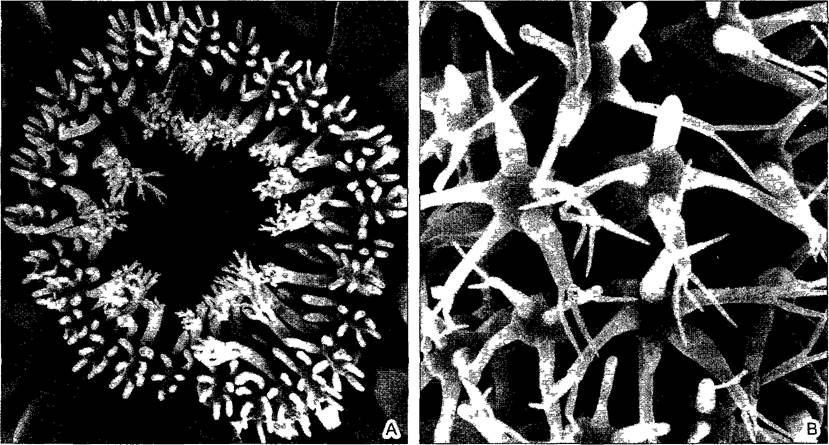
Рис. 4.1. Сложные структурные образования у сифональных водорослей выросты на гигантских клетках Dasycladaceae (ср рис 11 92) (СЭМ-фотография S Berger)
А—зонтиковидная мутовка ветвей Acetabufana polyphysoides, В—мутовки веточек Chlorocladus australasicus
Макроморфология крупных и потому легко наблюдаемых кормофитов долгое время была единственной основой систематики и таксономии. Однако видимое лишь кажется понятным, и чтобы постигнуть проблему, необходима работа рассудка. В связи с этим вспомним А Шопенгауэра «Задача состоит не столько в том, чтобы увидеть то, что прежде никто не видел, сколько в том, чтобы в увиденном каждым распознать то, что никто не заметил». Вот изречение другого классика, И -В Гете, одного из основателей сравнительной морфологии, открывшего межчелюстную кость у человека (1784) «Что сложнее всего? То, что тебе кажется наиболее легким видеть глазами, что лежит перед тобой» («Метаморфозы растений», 1790).
В настоящее время исследование причинных взаимодействии в развитии многоклеточных организмов (см гл 7) происходит чрезвычайно быстрыми темпами благодаря успехам молекулярной биологии. Ранее было возможно лишь очень ограниченное понимание морфогенеза, так как задействованные в нем сигнальные вещества и рецепторы появлялись на короткое время и в крайне низких концентрациях. Более доступно было изучение связей между структурами/функциями и приспособлениями организмов к окружающей среде и особым условиям жизни. формы организмов возможно понять в рамках телеономического рассмотрения исходя из их биологического значения и целесообразности.
Этот принцип введен в ботанику более 100 лет назад в двух эпохальных, хотя первоначально и вызвавших большую полемику трудах «Механический принцип в анатомическом строении однодольных» С Швенденера (1874) и «Физиологическая анатомия растении» Г Габерландта (1884) С тех пор ткани растений, например, определяли не только по их структуре, но и по функциям (см гл 3) Выяснение функционального значения особенностей строения организма и ранее было ведущей темой наблюдений ученых (см работу К К Шпренгеля «Раскрытая тайна природы в строении и оплодотворении цветка», 1793) Но только эволюционная теория Ч. Дарвина подвела научную основу под это направление исследований Конечно, роль принципа экономической целесообразности в интерпретации строения организмов нередко преувеличивали Особенно часто возникали ошибки из-за неверного представления о том, что естественный отбор сохраняет исключительно целесообразное и что поэтому живой мир должен быть устроен в высшей степени рационально и целесообразно В действительности отбор допускает недолгое выживание и нецелесообразного — и это совсем иное понимание Отбору как ограничительному принципу эволюции противостоят энтропийные процессы случайных наследственных изменении (мутации, рекомбинации, горизонтальный перенос генов) и симбиогенез. Они обусловили огромное видовое разнообразие и физиологическое, экологическое и морфологическое многообразие живого
Специфически морфологическим методом является типологический. Типы выявляют путем сравнения разных форм. Даже для больших систематических групп (родов, семейств и т.д.) при всей изменчивости и разнообразии входящих в них форм возможно определить константные и основополагающие особенности организации, которые и составляют тип соответствующей систематической единицы.
Еще И.-В. Гёте в этом же смысле использовал понятие «прообраз», впоследствии замененный понятием «план строения». Однако последнее слишком антропоморфно, что легко приводит к недоразумениям. Согласно В.Троллю, классику типологической морфологии предыдущего столетия, тип какой-либо группы организмов «можно выявить, но не показать». Тип представляет собой мысленный конструкт, абстракцию, основанную на выявлении общего, т.е. сходного в различных существах. Типологическая морфология не зависит от причинных и финалистских рассуждений. Она является основой составления «естественных» систем в биологии. Сама возможность выделения каких-либо морфотипов есть выражение иерархического филетического развития («родословных деревьев»). Показательно высказывание Ч. Дарвина о том, что морфология всегда означает вопрос о типе.
Для того чтобы описать все ныне живущие и сохранившиеся в ископаемом виде организмы, систематизировать и правильно называть их, разработана обширная терминология. Например, объяснения к рис. 4.2 дают нам представления о терминах, применяемых в описании листовых пластинок и их края. Определители растений содержат краткие сводки такой «прикладной морфологии».
Рис. 4.2. Некоторые формы края листа и листовой пластинки
Край листа А—цельнокрайный (кукуруза, здесь, как почти у всех однодольных, цельнокрайность сочетается с параллельным жилкованием — проводящие пучки листа проходят параллельно его краю, 2,8х), В—цельнокрайность у двудольного травянистого многолетника гречишки японской (Reynoutna japónica) с сетчатым жилкованием листьев (параллельное жилкование не обязательно для цельно- крайности, 2,8х), С—городчатый (хрен деревенский, 2,8х), D—зубчатый (каштан посевной, натур величина), Е—пильчатый (крапива, 1,7х), F—дважды пильчатый (Кеrriа, 1,5х), G—струговидный (одуванчик, Taraxacum officinale, 0,7х) Форма листа Н—лопастной лист (дуб черешчатый, Quercos robur, натур величина), I—перистосложный (рябина обыкновенная, Sorbus аucuparia, натур величина), К—пальчатолопастной лист (клен полевой, Acer campestre, 0,75х), L—пальчатосложный лист (лапчатка ползучая, Potentilla reptans, 0,75х)

Понятие «анатомия» (от греч. аnatemnein — резать, расчленять) имеет в ботанике несколько иной смысл, чем в медицине и зоологии. Чтобы увидеть скрытые в теле органы человека и животных, тела приходится разрезать в прямом смысле слова, чего совершенно не требуется для наблюдения поверхностно располагающихся органов большинства растений. Соответственно под анатомией растений понимают микроскопическое исследование расположения тканей в основных органах. Анатомия и макроморфология (органография) растений тесно взаимосвязаны, и поэтому в этой главе рассмотрены совместно.
4.1.1. Гомология и аналогия
Сходство не всегда означает одинаковый морфогенез или эволюционное родство. Наряду со сходствами, проистекающими от принадлежности к одному и тому же типу строения и тем самым — от родственных отношений (гомология), существуют сходства, обусловленные выполнением одних и тех же функций (аналогия). Гомология означает равнозначность зачатков, выражение сходной генетической информации, а аналогия, напротив, — выполнение одинаковых функций.
Например, в растительном и животном мире приспособления для воздушного полета возникали многократно и независимо. Все они основаны не на простой способности парить, а на использовании аэродинамических законов и образовании крыльев. Поэтому все крылья (насекомых, летучих рыб, птиц, летучих мышей и т.п.), крыловидные выросты плодов клена или семян видов рода Zanonia (см. рис. 11.194, D), равно как крылья самолетов и лопасти пропеллеров других летающих объектов, принципиально сходны, не будучи гомологичными друг другу. Обтекаемая форма быстро плавающих существ (и судов) также служит одной цели — уменьшить сопротивление среды. Такая форма свойственна некоторым бражникам, гаметам или даже огромным китам (аналогичное внешнее сходство которых с рыбами делает их «киторыбой»). В обиходной речи часто путают колючки и шипы (см. рис. 4.7), сходство которых обусловлено одинаковыми функциями (см. раздел 3.2.2.1).
Таким образом, если одинаковые потребности могут обусловливать сходство разных органов, то и, наоборот, гомологичные структуры становятся непохожими друг на друга из-за выполнения разных функций или приспособления к разным условиям внешней среды. Иллюстрацией тому служит различный вид одинаковых органов у одного и того же организма, например, морфологическая вариабельность листьев на разных частях растений (рис. 4.5,4.6; см, разделы 4.3.2, 4.3.3). Можно упомянуть также нетипичное функционирование/строение органов. Так, у некоторых растений боковые побеги с ограниченным ростом берут на себя функцию листьев (филлокладии; от греч. рhyllon — лист, klados — ветвь; рис. 4.3). Они имеют вид листьев, т.е. являются аналогом листа, но в действительности представляют собой укороченные побеги (гомологичны побегу). Побеговая природа филлокладиев проявляется в том, что, в отличие от настоящих листьев, они располагаются в пазухах чешуевидных или преобразованных в колючки листьев и могут нести цветки. У других растений роль листьев могут брать на себя воздушные корни (рис. 4.4). В этом случае они совсем не похожи на корни, но уплощенные и зеленые, как листья, вследствие наличия в их клетках хлоропластов. Такие превращения органов, обусловленные выполнением особых функций, в морфологии растений называются метаморфозами (от греч. — превращения; в зоологии этот термин имеет совершенно иное значение).
Рис. 4.3. Филлокладии — уплощенные побеги в качестве «листьев» (фото W. Barthlott).
Ветвь иглицы Ruscus aculeatus с похожими на листья, несущими цветки боковыми побегами, вырастающими из пазух чешуйчатых листьев (натур. величина; ср. рис. 4.34)

Рис. 4.4. Taeniophyllum zoliigeri — одна из орхидей, живущая как эпифит на дереревьях, с зелеными, лентовидными воздушными корнями (w), которые служат ассимилирующими органами (0,5х) (по К. Goebel)

Особенно важно четко различать гомологию и аналогию при изучении родственных связей между организмами (см. раздел 11.1.3). Эволюционное, или филетическое (от греч. phylon— ствол), родство проявляется только в гомологическом сходстве, т.е. в принадлежности к одному и тому же морфологическому типу. Есть разные (молекулярные, кариологические, морфологические и физиологические) критерии гомологии. Среди морфологических критериев самый основной — критерий положения: один орган гомологичен другому в том случае, если он занимает такое же пространственное положение в сопоставимых структурах. Как уже упоминалось, филлокладии находятся в пазухах кроющих листьев, что «типично» для боковых побегов (см. рис. 4.3). Еще один морфологический критерий — критерий переходных форм, связывающий резко различающиеся формы. Так, существуют промежуточные формы между катафиллами (редуцированными низовыми листьями), стеблевыми листьями, гипсофиллами (прицветными листьями), между листочками околоцветника и тычинками (см. рис. 4.6; 11.170), наконец, между листьями и листовыми колючками; все эти непохожие структуры представляют собой листья. В филогенетике такое же значение имеют ископаемые промежуточные формы между представителями разных систематических групп, в ходе эволюции утратившими сходство друг с другом. Наконец, для доказательства гомологичности органов особенно важно изучение ранних стадий их развития в онтогенезе (индивидуальном развитии организмов). Большинство органов в развитом состоянии выполняют определенные функции и обладают соответственно специфическими особенностями, тогда как сходство их зачатков еще позволяет распознать их гомологичность.
Рис. 4.5. Листовая серия морозника вонючего (Helleborus foetidusi 0,25х) (по D. von Denffer): А—семядоля; В, С—ювенильные листья; D—лист первого года развития; Е— стоповидно рассеченный лист второго года; F—переходный лист; G—I — верховые листья третьего года развития; К—лист обвертки цветка
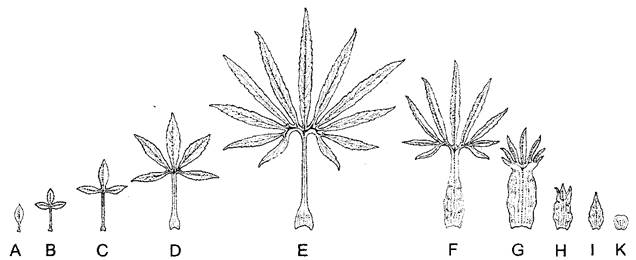
Рис. 4.6. Переходные формы между различными листовыми органами у розы собачьей, или шиповника (Rosa canina): А—наружные чашелистики 1 и 2 еще сохраняют перистость (сходство с обычными листьями), внутренние 4 и 5 цельные; чашелистик 3 перистый только с одной стороны, именно обращенной к чашелистику 2 (натур, величина); В—промежуточные формы между лепестками и тычинками. Стрелки — пыльники на краях лепестков (1,3х)

Дивергентная в целом эволюция, увеличивающая различия между формами, может приводить к сходству по отдельным признакам в результате адаптации к одинаковым условиям. Зоолог В.Хенниг (W. Неnnig) в рамках последовательно филогенетической систематики (кладизма) преобразовал терминологию, касающуюся понятий «аналогия/гомология». Эта кладистическая терминология стала общепринятой. Аналогия, т.е. сходство органов различного происхождения, названа ученым
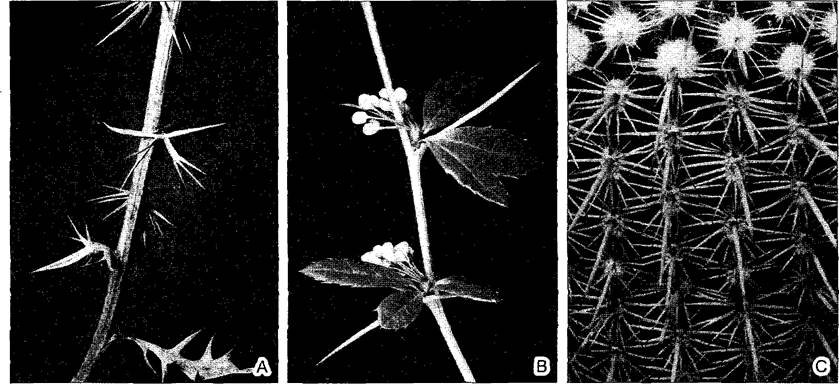
гомоплазией. В ней следует различать конвергенцию и параллелизм. Конвергенция, по Хеннигу, означает сходную форму негомологичных органов. Ее примерами могут служить колючки (рис. 4.7; см. рис. 4.36) и усики (см. рис. 4.69), т.е. органы, соответствующие листьям или метаморфизированным побегам. Под параллелизмом, напротив, понимают филетически независимое сходное преобразование гомологичных структур в разных систематических группах, например, появление стеблевых суккулентов в различных семействах.
Рис. 4.7. Превращение листьев в колючки: А, В—барбарис обыкновенный (Berbris vulgaris): А—прогрессирующее преобразование обычных листьев в листовые колючки в нижней части ветви (0,6х); В—из пазух полностью превращенных в колючки листьев вырастает укороченный побег, который в первый год образует зубчатые листья, а на второй год — цветки (0,9х); С—у большинства кактусов (в качестве примера приведен Notocactus rutilans) листья, также располагающиеся на укороченных пазушных побегах (ареолах), превращены в одревесневшие колючки. Функцию листьев выполняет зеленый, суккулентный стебель (1,9х)
4.1.2. Побег (кормус) и таллом
Всем папоротникообразным и голосеменным растениям свойствен общий морфотип, состоящий из трех основных органов — стебля, листа и корня. Такой тип организации растений называют кормусным (от греч. коrmos — ствол, побег). Взаимное расположение осевых органов у кормофитов (побеговых растений) всегда одинаковое: листья неизменно располагаются на стеблях и никогда не бывают на корнях. Корни образуют эндогенные боковые корни, а побеги — совершенно иные, экзогенные, боковые побеги (ветвление); однако придаточные корни могут возникать на побеге, а придаточные побеги — на корне. Следует отметить, что цветок не является основным органом; это лишь укороченный побег, несущий спорофиллы и служащий для воспроизведения растений.
Не гомологичны кормусу вегетативные тела многоклеточных водорослей, а также печеночников1. Их тела называют талломами (от греч. thallos — листва). Обзор их морфологической организации дан в гл. 5. Настоящая глава посвящена морфологии и анатомии кормофитов как наиболее известной, изученной, многочисленной, научно значимой и эволюционно самой молодой и продвинутой группе растений.
1 Большинство видов печеночников имеет побеговую организацию. — Примеч. ред.
В функциональном отношении все живые существа — высоко упорядоченные образования. Обычно этот абстрактный функциональный порядок явным образом проявляется в упорядоченности структур, ниже приведены примеры этою Закономерное повторение одних и тех же или сходных структурных элементов называют симметрией (от греч symmetros — соразмерность)
Есть три основные формы симметрии:
• Метамерия — поступательная симметрия — повторение сходных элементов вдоль оси на одинаковом расстоянии и в той же ориентации гомономная метамерия (рис А)
Рис. А. Примеры метамерии у растений.
А—перистый лист Potentilla anserine (натур величина), В—часть перышка первого порядка вайи щитовника мужского Dryopteris filix-mas (нижняя сторона с сорусами, под светлыми почковидными индузиями (покрывальцами) скрываются пучки созревших спорангиев) (2,5х), С—расположение продольными рядами ареол (укороченных побегов с листовыми колючками) кактуса цереуса, Сеreus pasacana (0,2х), D—кроющие чешуи еловой шишки (1,5х) Другие примеры метамерии представляют хвощи (см рис 11 141, Е, К)

Особые формы получаются при непрямых осях (кривая, спираль) и полярной оси, вдоль которой элементы, например, становятся все меньше (гетерономная метамерия). Прямая или изогнутая ось метамерии может стать полярной (вектором), если элементы вдоль нее одинаково асимметричны. Это имеет место у многих биополимеров. Благодаря полярности осей обычно устанавливается направление синтеза (5' —» 3' у нуклеиновых кислот, N-конец —» С-конец у полипептидов; удлинение с плюс-конца у микротрубочек или актиновых микрофиламентов). Число элементов симметрии при гомономной метамерии не ограничено. В определенных случаях реально достигаются очень высокие числа, например, нуклеотидов в молекулах ДНК.
• Радиальная симметрия — симметрия вращения — повторение сходных одинаково ориентированных элементов под равными углами вокруг оси симметрии (рис. В). Число элементов симметрии ограничено, ось симметрии можно характеризовать этим числом.
Рис. В. Примеры радиальной симметрии
А—расположение корзинок у мордовника Echinops sphaerocephalus (0,8х) Много других примеров шаровидной симметрии имеется у плодов и семян, В—Primula, садовая форма (чем объясняется шестимерность этого цветка, дикорастущие формы имеют 5-мерный цветок) (1,2х), С—вращательная симметрия 5-го порядка цветка олеандра (0,8х)

• Билатеральная симметрия — зеркальная симметрия (рис. С) — отражение в плоскости симметрии — медиане: есть два элемента симметрии — структура и ее отражение. Этот тип симметрии господствует в мире животных и представлен в нашем собственном теле. Но и у растений встречаются бесчисленные примеры билатеральной симметрии: большинство листьев и так называемые зигоморфные цветки. Радиальная симметрия путем определенных преобразований поперек оси симметрии переходит в билатеральную. Соответственно зеркальная симметрия встречается у организмов преимущественно в тех случаях, когда два определяющих форму вектора перекрещиваются (у животных: сила тяжести/направление движения; у растений: сила тяжести/направление роста — поэтому прежде всего у органов, отходящих вбок от вертикальной оси). Билатеральная симметрия почти всегда связана с дорсовентральностью, т.е. с различиями между верхней и нижней сторонами (лат. dorsum — спина; venter — живот, брюхо).
Рис. С. Билатеральная симметрия (В—фото К. и Н. Rasbach; С—фото W. Ваrthlott): А, В—цветки орхидей, как у многих других растений, зигоморфны. В качестве примера приведены цветки Венерина башмачка (Раphiopedilum—А, 0,5х) и офриса насекомоносного (Оphysinsectifera—В, 2х), цветки офриса самцы определенных насекомых принимают за самок, при попытках спаривания они переносят поллинарии с одних цветков на другие (ср. рис. 11.227) и таким образом осуществляют опыление. Эти цветки не только билатерально симметричны, но и дорсовентральны (отличия верх/низ или перед/зад). Билатеральны и дорсовентральны также подавляющее большинство типичных листьев: С—лист тропической Веrtouvelia houtteana из Melastomataceaе; D—зеркально симметричны и некоторые одноклеточные, например, декоративная водоросль Micrastenas radiata (190х)

Комплексная симметрия проявляется в тех случаях, когда комбинируются два или все три основных типа симметрии, то есть варианты разных типов симметрии перекрываются в одной структуре (рис. D). Простой пример такой симметрии — мутовчатое расположение листьев: отдельные мутовки радиально симметричны, но наряду с этим проявляется продольная метамерия побега, причем листья соседних мутовок чередуются (см. 4.2.2). В таких случаях симметрия особенно отчетливо показывает объединение элементов в систему.
Рис. D. Примеры комплексной симметрии
А—цветок белозора болотного (Parnassia palustris), венчик цветка 5-членный, как и снабженные железистыми головками стаминодии (тычинки, преобразованные в нектарники, здесь ложные, так как нектар не содержит сахара) и обычные тычинки, пестик, напротив, 4-членный (1,3х), В—цветок страстоцвета Passiflora caerulea с многочисленными нитевидными радиально симметрично расположенными выростами цветоложа, 5 точно так же радиально расположенными тычинками и 3 плодолистиками, 3 темных рыльца ограничивают два более крупных и один меньший углы так, что расположение рылец становится билатерально симметричным (0,6х), С—часть ветки с плодами Phytolacca, зигоморфные плодолистики собраны в радиально симметричное образование, гинецей цветка Цветки, как и плоды, расположены на горизонтальных ножках, которые в свою очередь отходят от вертикальной главной оси по спирали (метамерия) (1,3х)

Временную метамерию — регулярные ритмы — возможно представить на плоскости в виде диаграммы, показав время как ось; ее симметрия станет очень наглядной. У живых существ можно найти бесчисленные примеры ритмических процессов, наряду с движениями (биение жгутиков, взмахи плавников и крыльев, шаг и бег) также эндогенные ритмы (см. 7.7.2.3) и периодически повторяющийся морфогенез, например, побега, пространственно выражающийся в его метамерии. В молекулярной области ему соответствуют ритмично повторяющиеся реакции синтеза при образовании макромолекул.
Правильные узоры получаются в тех случаях, когда идентичные одинаково ориентированные элементы тесно сближены. Круги или шары одного размера при плотнейшей упаковке образуют гексагональный узор (рис. 4.17): каждый элемент окружен шестью другими, и имеется три выделенных направления, пересекающиеся под углами соответственно 60° или 120°. Однако в большинстве биологических узоров отдельные элементы не вполне идентичны. Они, например, варьируют по величине, ориентированы не абсолютно одинаково, а расстояния между ними не равны, а варьируют в определенных пределах (рис. Е, F). Тем не менее они ясно отличаются от хаотичных узоров, в ориентации и расположении элементов которых невозможно уловить никаких закономерностей.
Рис. Е. Статистические и регулярные узоры
А—в черном хлебе «здоровье» запеченные зерна пшеницы образуют случайный узор с нерегулярными расстояниями между (светлыми) зернами (0,5х), В—зерна кукурузного початка при почти одинаковой величине и плотности расположения отчетливо демонстрируют регулярное расположение, распределение темных зерен — результат менделевского расщепления 1 3—из-за случайностей мейоза создает случайный узор (0,5х), С, D—в этих примерах высокой, но не совершенной регулярности расстояния между соседними элементами узора сходны, но не идентичны С—устьица на верхней поверхности плавающего листа кувшинки Nymphaea alba (70х), D— нижняя сторона шляпки трутовика (Polyporus, 1,3х)

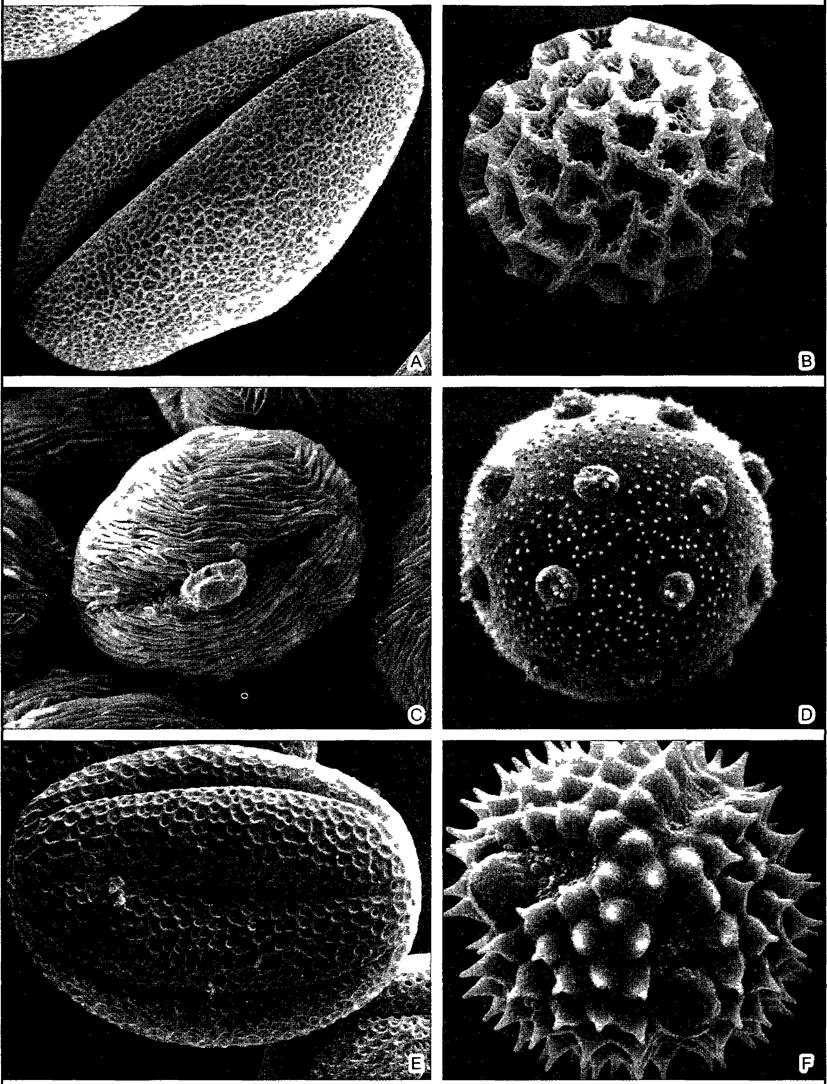
Рис. F. Узоры на отдельных клетках поверхностные структуры пыльцевых зерен (ср также рис 1 176 и 11 214) (препараты и СЭМ-фотографии T Esche)
А—Stachys recta, В—Phlox (садовая форма), С—Centaunum erythraea, D—Silene nutans, E—Thymus pulegioides, F—Aster Imosyris
Неполная симметрия биологических узоров связана с тем, что жесткая упорядоченность не допускала бы никаких проявлений жизни: предпосылкой всех процессов развития, биосинтеза и движения являются нарушения симметрии. И в эволюции организмов неоднократно происходят роковые нарушения симметрии. Знаменательно, что кристалловидные вирусные частицы в клетках хозяина требуются для облегчения размножения вирусов.
Особое значение в биологии имеет комплементарная симметрия (антисимметрия): два неодинаковых, но гармонирующих друг с другом структурных элемента объединяются для выполнения одной функции. Известные примеры из техники — замок и ключ или розетка и штепсельная вилка; из мира животных — суставы, совокупительные органы. Антисимметричные молекулярные структуры часто осуществляют процессы узнавания или размножения: фермент/субстрат; рецептор/лиганд; транслокатор/переносимое вещество, антиген/антитело и т.д. Антисимметричны комплементарные основания полинуклеотидных цепей двойной спирали ДНК, а также кодон и антикодон при трансляции. Самоорганизация надмолекулярных биоструктур, например, четвертичная структура белков или вирусные частицы — вся без исключения основана на комплементарной симметрии молекул и их частей.
В антисимметричных структурах строение одного элемента комплементарно строению другого. Тем самым исключено непосредственное сходство элементов. В сложных функциональных системах дело доходит до того, что все их элементы различны, но при этом комплементарно симметричны. Функциональная связь в таких случаях часто появляется только в особенностях взаимного положения, но никак не в морфологии элементов. Чем больше элементов в такой функциональной системе, тем разнообразнее ее действия и тем ниже в целом степень ее симметрии в морфологическом отношении. С этим связана, например, низкая симметрия многоклеточных структур. Крайний пример представляют амебоидные клетки. Однако несимметричные организмы встречаются значительно реже, чем симметричные, которым, по-видимому, благоприятствовал отбор. Это и понятно: симметрия означает (также) повторение; для развития и функционирования симметричной системы необходимо значительно меньше информации, чем несимметричной.