БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
6. ФИЗИОЛОГИЯ ОБМЕНА ВЕЩЕСТВ
6.15. Синтез тетрапирролов
Как тетрапирролы с кольцевой молекулой (порфирины), так и нециклические тетрапирролы выполняют в растениях различные функции. Хлорофиллы и бактерио- хлорофиллы поглощают световую энергию в процессе фотосинтеза (см. 6.4.2); гем является составной частью цитохрома, каталаз и пероксидаз и обнаруживается в леггемоглобине корневых клубеньков. Цитохромы являются переносчиками электронов, например, в процессе клеточного дыхания (см. 6.10.3) или фотосинтеза (см. 6.4.2), цитохром Р450 — составная часть монооксигеназ (см. 7.6.3.2), гем каталаз отвечает за устранение активного кислорода в форме Н2O2, в пероксисомах и глиоксисомах (см. 6.5.6; 6.12). Пероксидазы выполняют многочисленные функции в окислительных реакциях: например, они служат для обезвреживания ксенобиотиков (потенциальных ядов) и важны для образования лигнина (см. 6.17.2). Сирогем в качестве составляющей сульфитреду ктазы (см. 6.7) и нитритредуктазы (см. 6.6.1) тоже является переносчиком электронов. Леггемоглобин корневых клубеньков бобовых (см. 9.2.1) служит для запасания молекулярного кислорода при фиксации атмосферного азота.
В отличие от железосодержащих гемов и содержащих магний хлорофиллов, тетрапиррольное кольцо витамина В12 (цианкобаламина) имеет в качестве центрального атома кобальт. Витамин В12 синтезируется только несколькими видами бактерий. Он не синтезируется растениями, которые его не используют, и животными, которым он необходим. Животные и человек потребляют витамин В12 с животной пищей или получают его благодаря кишечной микрофлоре (растительноядные животные!). Потребность человека в витамине В12 составляет всего лишь несколько микрограммов в день, однако при строгом вегетарианском питании у людей могут появиться признаки авитаминоза (анемия, малокровие).
Нециклические тетрапирролы являются хромофорными группами фикобилипротеидов — дополнительных пигментов фотосинтеза цианобактерий и красных водорослей (см. 6.4.2; 6.4.3). Близкий по строению к фикоцианобилину и фикоэритробилину фитохромобилин представляет собой хромофор растительных рецепторов красной части спектра (фитохромы — см. 7.7.2.4).
Биосинтез порфириновой системы происходит у зеленых растений в пластидах, которые в дальнейшем также осуществляют начало синтеза хлорофиллов, гема и сирогема. Митохондриальный синтез гема идет на основе предшественников, образованных в пластидах. Предположительно, пластиды экспортируют гем для его использования в качестве простетической группы гемсодержащих ферментов других компартментов. Синтез тетрапирролов на рис. 6.112 показан максимально полно (насколько он изучен на данный момент).
Рис. 6.112. Компартментация и регуляция метаболизма тетрапирролов в растении. Активизация протохлорофиллидоксидоредуктазы (ПОР А) светом типична для покрытосеменных растений, которые зеленеют только на свету

Сложные биохимические превращения тетрапирролов здесь можно представить только в общих чертах (рис. 6.113). Основная составная часть системы тетрапирролов, порфобилиноген, появляется в результате конденсации двух молекул δ-аминолевулиновой кислоты. Кислота возникает у растений, цианобактерий и многих других бактерий из глутамата (у животных, дрожжей и некоторых бактерий из сукцинил-КоА и глицина), восстанавленного до 1-семиальдегида, который превращается в δ-аминолевулиновую кислоту посредством внутримолекулярного тиаминпиро- фосфатзависимого трансаминирования. Интересно, что в качестве активирующего предварительного этапа для восстановления карбоксильной группы выступает не остаток фосфорной кислоты, а глута- мил-тРНК (вовремя пластидной трансляции она служит также донором глутамата); восстановление происходит на этапе аминоацил-тРНК.
Рис. 6.113. Биосинтез тетрапирролов из глутамата. Соединения на этапах до уропорфириногена являются собственно тетрапирролами, протопорфирин IX— общий предшественник хлорофилла и гема (см. рис. 6.112), как и нециклические тетрапир- ролы, которые возникают из протогема посредством размыкания кольца
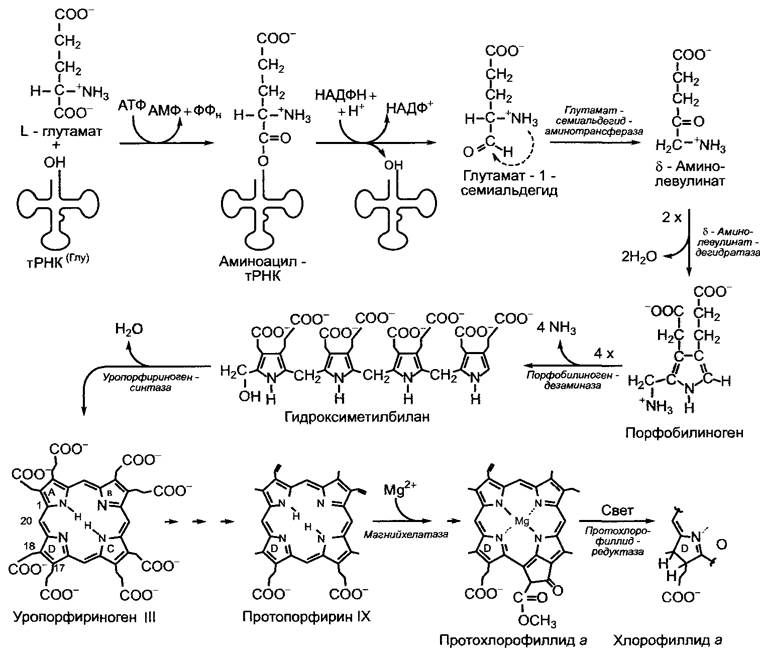
Четыре молекулы порфобилиногенов посредством дезаминирования конденсируются в нециклическую молекулу гидроксиметилбилана (предварительный этап), который циклизуется в первый циклический тетрапиррол — уропорфириноген III в результате воздействия уропорфирино- генсинтазы и отщепления воды. Через несколько промежуточных этапов синтезируется протопорфирин IX, который преобразуется в протогем при помощи феррохелатазы или в протохлорофиллид а под воздействием магнийхелатазы. Восстановление D-кольца приводит к превращению протохлорофиллида а в хлорофиллид а, на который хлорофиллсинтетаза, называемая также пренилтрансферазой, переносит остаток фитола (структура хлорофилла — см. рис. 6.44). Хлорофилл b образуется из хлорофилла а или хлорофиллида а, детали неизвестны. Синтез сирогема отделяется уже на этапе уропорфириногена III. Образование нециклических тетрапирролов осуществляется размыканием кольца на предварительном этапе синтеза порфирина, который возникает из протогема (см. рис. 6.112).
Большинство зеленых водорослей, голосеменных, фотосинтезирующих бактерий и цианобактерий синтезируют хлорофилл на свету и в темноте, покрытосеменные — только на свету. У них протохлорофиллидредуктаза регулируется светом.
Регуляция синтеза тетрапирролов в соответствии с потребностью растения осуществляется следующим образом (см. рис. 6.112). Так, конечные продукты протохлорофиллид и протогем ингибируют синтез δ-аминолевулиновой кислоты, магнийхелатаза ингибируется протохлорофиллидом и хлорофиллидом. Образование δ-аминолевулиновой кислоты активируется на свету при помощи фитохрома. Благодаря этим механизмам предотвращается избыточное накопление в темноте фотореактивных молекул протохлорофиллида.