БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
8. ФИЗИОЛОГИЯ ДВИЖЕНИЙ
8.4. Прочие движения
В растительном царстве распространены гигроскопические и когезионные движения. Гигроскопические движения основываются на неравномерном набухании и протекают без непосредственного участия живых клеток. Они служат для распространения спор, пыльцы, семян и плодов. Движение обусловлено чисто физическими причинами и основывается на различиях в растяжении или соответственно сокращении фибриллярных слоев при набухании или при потере влаги. Характер набухания клеточной стенки определяется преобладающим направлением параллельных слоев микрофибрилл во вторичной клеточной стенке. Растяжение или соответственно сокращение происходит преимущественно перпендикулярно направлению микрофибрилл. Если слои ткани с различным направлением микрофибрилл и различным составом клеточных стенок лежат один над другим, то при изменяющейся влажности ткани (например, вследствие высыхания в процессе созревания и в зрелом состоянии при различном состоянии набухания в сухой или влажной среде) это приводит к скручиванию. Способность к набуханию основных компонентов клеточных стенок показана в следующем ряду в порядке возрастания: лигнин < целлюлоза < гемицеллюлоза < пектин.
Внешние зубцы перистома на спорангиях лиственных мхов, состоящие большей частью только из частей клеточных стенок двух граничащих друг с другом клеточных слоев, при высыхании гигроскопично изгибаются внутрь или наружу в зависимости от своей тонкой структуры и посредством этих движений, следующих за колебаниями влажности воздуха, содействуют или соответственно препятствуют рассеиванию спор. На рис. 8.36 представлен следующий пример: движение зубца перистома при высыхании определяется тем, что микрофибриллы во внешней пластинке расположены поперек продольной оси зубца, так что этот слой предпочтительно укорачивается по продольной оси. Внутренняя пластинка, напротив, вследствие расположения фибрилл вдоль оси только слегка сморщивается в толщину, не уменьшаясь по длине. Будучи прочно связанной с внешним слоем стенки, она из-за этого препятствует укорачиванию зубца и вызывает его скручивание наружу. Строение клеточных стенок перистомов одного рода мха очень многообразно, и вследствие этого направления движений различны — как приспособления в зависимости от конкретных экологических потребностей. Сходные гигроскопические движения свойственны и прицепкам спор Equisetum (см. 11.2; рис. 11.141, Н, J), и капиллициям некоторых слизевиков (см. 11.2; рис. 11.17, Е).
Многие плоды-коробочки открываются, когда протопласты клеток стенки плода отмирают и клеточные оболочки начинают высыхать (ксерохазия, например, у Saponaria), другие остаются закрытыми в высохшем состоянии и открываются лишь при смачивании (гигрохазия, например, у видов Mesembryanthemum, Sedum, Verónica). К анизотропному набуханию отдельных слоев чешуй сводятся также движения открывания (при высыхании) и закрывания кроющих чешуй шишек хвойных (например, у шишек сосны, см. 11.2; рис. 11.203).
У плодиков видов Erodium (рис. 8.36, В) при высыхании происходит спиральное закручивание. При повторном смачивании ости пытаются снова выпрямиться и, если их свободные концы наталкиваются на опору, вкручивают плодики в почву. Подобным образом ведут себя также ости зерновок некоторых злаков (например, Stipa). Гигроскопическую подвижность проявляют и волоски летучек многих семян и плодов (например, одуванчика).
Рис. 8.36. Гигроскопические движения: А — зубец внешнего перистома коробочки мха Orthotrichum diaphanum в сухом и набухшем состоянии. Показаны наружный и внешний слои клеток зубца со схематическим изображением направления укладки микрофибрилл. Рядом коробочка мха с открытым (слева) и закрытым (справа) перистомом (на схеме даны лишь два зубца перистома); В — плодик Erodium gruinum в сухом и набухшем состояниях

У североафриканского растения из крестоцветных Anastatica hierochuntica («иерихонская роза») сухие ветви свертываются внутрь, а увлажненные широко раскидываются. Считалось, что сухие шаровидные растения Anastatica могут перекатываться ветром и таким образом распространять семена, но это мнение не подтвердилось.
В отличие от гигроскопических движений когезионные движения основаны на весьма значительных силах сцепления молекул воды даже в тонких водных слоях (см. 6.3.2.2, уравнение 6.34).
Так, например, у отдельных клеток кольца дугообразно охватывающего спорангий папоротника (рис. 8.37) внутренние и боковые стенки утолщены в отличие от внешних стенок. При созревании спорангия эти клетки начинают медленно терять воду. Но поскольку вода, пропитывающая клеточные стенки, прочно удерживается ими, и вода, заполняющая клетку, вследствие значительных сил сцепления между ее молекулами также сначала не отделяется от нее (для этого требуется гидростатическое давление отрицательнее -25 МПа!, см. 6.3.5), то по мере испарения внутриклеточной воды верхние части антиклинальных клеточных стенок сближаются из-за прогибания внутрь тонких внешних стенок. В результате на верхней части спорангия возникает тангентальное натяжение. Вследствие этого две клетки в заранее определенном месте (стомий) расходятся, и мертвая стенка спорангия, начиная с этого места, медленно разрывается, отгибаясь наружу. Когда деформация дугообразно сжатых клеток достигает такого предела, что силы сцепления внутриклеточной воды в них последовательно одна за другой преодолеваются, го в отдельных клетках кольца происходит компенсация напряжений. Каждый такой «скачок» в клетке вызывает рывок; в результате их совокупности отогнутая назад стенка спорангия, «сотрясаясь», возвращается в исходное положение, при этом рассыпая споры. На очень похожем механизме основано открывание пыльников, где лежащие в стенке пыльника волокнистые клетки эндоте- ция на основании жесткости их стенок функционируют подобно клеткам кольца. В перегородках спорангиев и у элатер многих печеночных мхов действуют похожие механизмы (рис. 8.38).
Рис. 8.37. Когезионный механизм у кольца спорангия Dryoptens: А — еще закрытый спорангий; В — разрывание (клетки вследствие когезионного сцепления воды сжимаются, кольцо изгибается наружу); С — конечное состояние после повторного сжатия кольца (напряжение снято вследствие проникновения пузырьков воздуха)
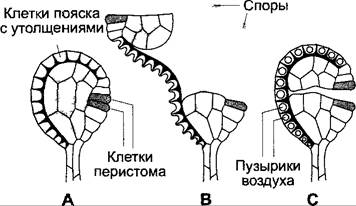
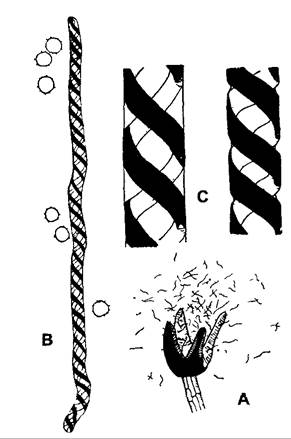
Рис. 8.38. Элатеры печеночного мха Сephalozia biicuspidata: А — вскрывшаяся коробочка (6х); В — отдельная элатера со спорами (100х); С — часть элатеры; слева — насыщенной водой, справа — после частичного испарения наполняющей воды (425х)
Ловчий механизм пузырьков Urticularia (см. бокс 4.4, В) также основан на действии когезионных сил заполняющей клетки воды. Вследствие активного выхода ионов N+. К+ и СI- из заполняющей клетку воды через стенку пузырька наружу и следующего за ним осмотического выхода воды пузырек теряет до 40 % воды. Таким образом образуется отрицательное гидростатическое давление по сравнению с окружающей средой, что заметно по вогнутости готового к поимке пузырька-ловушки. Прикосновение к щетинке-антенне вызывает открывание крышки-клапана, и окружающая вода вместе с жертвой засасывается в ловушку («принцип глотательной ловушки»).