БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
10. ЭВОЛЮЦИЯ
10.2. Типы и причины естественной изменчивости
10.2.1. Естественный отбор
Наблюдаемые в природе примеры внутривидовой генетической изменчивости чаще всего не случайны, а коррелируют с особенностями окружающей среды и свойствами исследуемых видов растений. Уже с начала XVIII в. была известна внутривидовая изменчивость, например, сроков цветения лесных деревьев северного и южного происхождения. Однако до экспериментов шведского ботаника Г.Турессона в 20-е годы XX столетия не было единого мнения, имеет ли такая внутривидовая изменчивость генетическую основу или она в большей мере результат условий местообитания. Культивируя при одинаковых условиях растения различного происхождения, относящиеся к широко распространенным и встречающимся в разных экологических условиях видам, Турессон смог показать, что наблюдаемые в естественных местообитаниях различия в большинстве случаев хотя бы частично сохраняются в культуре. Из этого он сделал вывод, что внутри вида существует генетическая изменчивость изучаемых признаков. Так как материал из различных мест со сходными экологическими условиями (рис. 10.25) (например, дюнные популяции ястребинки зонтичной Hieracium umbellatum) всегда демонстрировал сходные признаки и отличался от материала из мест с другими условиями (например, из популяций на прибрежных скалах), Турессон установил, что внутривидовая генетическая изменчивость коррелирует с условиями местообитания. Данные и выводы ученого удалось подтвердить и уточнить в большом числе сравнительных исследований. Соответственно можно уверенно считать, что коррелирующая с условиями среды внутривидовая генетическая изменчивость оказывается очень распространенным явлением.
Рис. 10.25. Экологические расы калифорнийского тысячелистника (Achillea Ianulosa, тетраплоид) с различной высоты над уровнем моря (1 400 — 3 350 — 2 100 м) вдоль транссекты длиной около 60 км через Сьерра-Неваду и примыкающий район Большого Бассейна (примерно 38° с. ш.). Около 60 индивидов из каждой популяции были выращены из семян в Стэнфорде (30 м). Диаграммы показывают наследственную изменчивость высоты побега, среднее значение (стрелками) и рисунок типичного индивида из каждой популяции

Вдоль транссекты с запада на восток через Калифорнию от побережья до высокогорий калифорнийских гор Сьерра-Невада и пограничного с ней Большого Бассейна удалось зарегистрировать генетическую изменчивость многих видов, коррелирующую с климатическими факторами. Например, растения тысячелистника Achillea lanulosa, происходящие с различных высотных уровней хр. Сьерра-Невада, отличаются высотой от растений того же вида, произрастающих в Большом Бассейне (см. рис. 10.251. Исследование злака Andropogon scoparius, широко распространенного в Северной Америке, показало, что северным формам для стимуляции цветения требуется более длительное освещение (15 ч в день), чем растениям южного происхождения (14 ч в день). Для понимания этого наблюдения необходимо знать, что длина дня летом возрастает с широтой, а запуск цветения у многих растений зависит от длины дня. Корреляцию генетической изменчивости с различными свойствами почвы смогли показать у Achillea borealis. В эксперименте формы, взятые с серпентиновых почв, хорошо растут на серпентиновых почвах (серпентин — горная порода, на которой возникают богатые магнием и бедные известью почвы). Но формы того же вида, которые в природе не встречаются на серпентиновых почвах, в экспериментальной культуре на серпентиновых почвах растут плохо. Удалось показать, что растения Anthoxanthum odoratum из бедных известью местообитаний нуждаются для роста в значительно меньшем количестве извести, чем растения того же вида родом из местообитаний, богатых известью. Многочисленные виды (например, Agrostis capillaris, Anthoxanthum odoratum, Festuca ovina, Mimulus guttatus, Plantago lanceolata, Rumex acetosa) имеют генотипы, которые позволяют им расти на почвах, загрязненных тяжелыми металлами, например, кадмием, медью, цинком, в то время как другие их генотипы не в состоянии это обеспечить. Внутривидовая дифференциация может коррелировать не только с абиотическими факторами, но и с биотическими. Как и многие другие виды, клевер ползучий (Trifolium repens) содержит цианогенный гликозид, ферментативное расщепление которого, например, при повреждении тканей приводит к высвобождению синильной кислоты. Данный вид полиморфен по этому признаку. Можно показать, что относительная частота цианогенных (с цианогенным гликозидом) и ацианогенных (без этого гликозида) генотипов коррелирует не только с температурой местообитания (рис. 10.26) при мелкомасштабном рассмотрении, но и с частотой улиток, поедающих этот клевер (при крупномасштабном рассмотрении). Plantago lanceolata с сильно вытаптываемых местообитаний в отличие от подорожника из мест, где ходят мало, имеет не прямостоячую форму роста, а стелющуюся или приподнимающуюся. Внутривидовая дифференциация, коррелирующая с факторами окружающей среды кроме вышеназванных примеров показа
на также и в отношении доступного для фотосинтеза света, засухоустойчивости, морозоустойчивости, устойчивости к нападению паразитов и травоядных животных, привлечения опылителей, конкуренции с окружающими растениями и т. п. (рис. 10.27).
Рис. 10.26. Клинальная изменчивость у клевера ползучего (Тritolium гереns). Частота аллеля А, ответственного за образование цианогенного гликозида (альтернативный аллель а) в популяциях (кружки с черными (= А) и белыми (= а) секторами), коррелирует с январскими изотермами от Средиземноморья до Северной Европы (карта слева), в Альпах — с высотой над уровнем моря (круговые диаграммы справа)
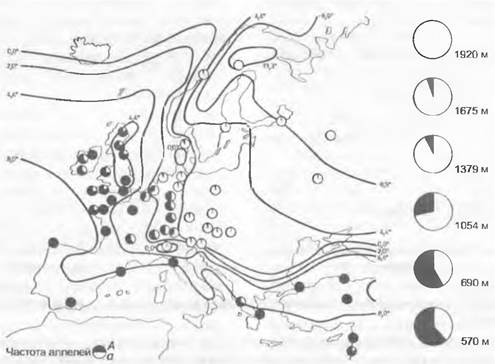
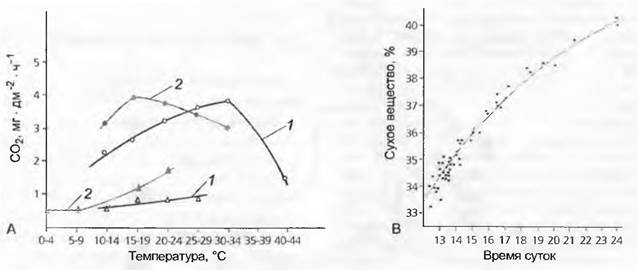
Рис. 10.27. Экологическая дифференциация у семенных расте: А — экотипы кисличника двухстолбчатого (Оxyria digyna, Polygonaceae) и их различные физиологические нормы реакции: средние показатели фотосинтеза (кружки) и дыхания (треугольники), измеренные в мг СО2 на квадратный дециметр поверхности листа, в зависимости от температуры у южной альпийской (1) и северной арктической (2) рас; В — клинальная изменчивость сосны обыкновенной (Pinus sylvestris): при одинаковых условиях выращивания 52 растения европейского происхождения показывают тесную корреляцию массы сухого вещества хвои (как меры устойчивости к холоду) со временем суток в первый весенний день (средняя температура +6 °С) в их естественном местообитании (как мера географической широты, континентаньности и длины вегетационного периода)
Детальное пространственное размещение генетических вариаций в природе может быть разных типов. Если коррелирующий с каким-либо признаком растения фактор окружающей среды распределен в природе прерывисто (к примеру, щелочные и кислые почвы над известковыми и силикатными породами в Альпах), растения могут показывать более или менее дискретную изменчивость. Если же, напротив, фактор среды изменяется непрерывно (например, снижение средних температур с увеличением высоты над уровнем моря), то и в исследуемом растительном материале надо ожидать непрерывной генетической изменчивости. Оба эти типа внутривидовой изменчивости давно уже различали как экотипическую (экотип) и экоклинальную (экоклина) изменчивость. Разграничение этих типов бывает затруднено тем, что анализируемый признак может коррелировать не с одним, а с двумя или несколькими факторами окружающей среды, изменяющимися в пространстве разнонаправленно.
Обнаруженные типы внутривидовой генетической изменчивости зависят и от многих экспериментальных параметров. Так, сравнение пространственно далеких популяций чаще всего показывает более четкие различия, чем сравнение соседних популяций, а сравнение выращенного из семян потомства представителей двух популяций будет демонстрировать менее выраженные различия, нежели сравнение взрослых растений, взятых из естественных местообитаний.
Естественный отбор общепризнан как важнейшая причина внутривидовой изменчивости, коррелирующей с факторами окружающей среды. Причиной естественного отбора является то, что разные индивидуумы (генотипы) популяции генетически различаются по успешности размножения в данной среде. Различия в успешности размножения проявляются в разной вероятности выживания до репродуктивного возраста, а после достижения репродуктивной фазы — в разном репродуктивном успехе. Разные шансы выжить и различный репродуктивный успех можно в совокупности назвать приспособленностью генотипа. Таким образом, объектом естественного отбора оказывается индивидуальный генотип, точнее, реализованный последним фенотип, т. е. индивидуум. Если разные генотипы популяции имеют разную приспособленность, то возникает естественный отбор. В результате естественного отбора может измениться относительная частота аллелей как на разных стадиях развития одного поколения, так и у последовательных поколений. Изменение частот аллелей в последовательности поколений можно назвать эволюцией. Конкретным (но мысленным) примером эволюции путем естественного отбора может служить один из видов тысячелистника — Achillea lanulosa, произрастающий вдоль транссекты через Калифорнию. Если две идентичные по генетическому составу популяции с генетически изменчивой высотой растений (с чем связаны число цветков и скорость развития до первого цветения) вырастить на побережье и в высокогорье, то их развитие в течение нескольких поколений, вероятно, пошло бы разными путями. В прибрежной популяции высокорослые растения с большим числом цветков дали бы в среднем больше потомков, чем низкорослые растения с немногими цветками. При более или менее постоянной численности популяции это приведет к увеличению относительной частоты высокорослых растений. В горах, где высокие растения из-за короткого вегетационного периода, возможно, даже не успеют зацвести или лишь самые первые их цветки смогут образовать семена, увеличилась бы относительная частота низкорослых растений. Таким образом, естественный отбор объясняет, как эволюцию, так и генетическое приспособление (адаптацию), ибо, по-видимому, рационально считать вариабельность высоты растений адаптацией к условиям произрастания. Итак, изменчивость, коррелирующую с окружающей средой, в большинстве случаев следует понимать, как адаптивную изменчивость.
Признак можно интерпретировать как генетическую адаптацию в тех случаях, если он наследуется и способствует повышению вероятности выживания или успеха в размножении, т. е. повышению приспособленности индивидуума. Термином «адаптация» обозначают как процесс приспособления, так и состояние приспособленности. Процесс генетической адаптации начинается с того, что какое-то свойство возникает благодаря мутации, т. е. совершенно случайно. Если новое свойство индивидуума повышает его приспособленность, то вторым шагом возникновения генетической адаптации становится распространение этого свойства в популяции благодаря естественному отбору.
Для доказательства роли естественного отбора как причины естественной генетической изменчивости недостаточно наблюдать корреляцию генетической изменчивости с определенной вариабельностью среды. Необходимо многократно показать, что разные генотипы в разных условиях среды имеют различную приспособленность или что генотип в своей естественной среде обитания, имеет более высокую приспособленность, чем в чуждой ему среде. Это можно осуществить,
либо подвергнув разные генотипы1 действию определенного решающего фактора среды и измерив их приспособленность, либо произведя в природе реципрокную пересадку материала различного происхождения. Наконец, можно определить различия приспособленности индивидуумов одной популяции в естественных условиях.
1 Все эксперименты возможны не непосредственно с генотипами, а с организмами, т. е. с фенотипами, обладающими разными генотипами. — Примеч. ред.
Первый вариант исследования был реализован на подорожнике большом (Plantago major). Отмечено, что растения в местах, где регулярно ходят, имеют более или менее распростертые цветоносы, а растения из мест, где вытаптывания нет, имеют прямостоячие цветоносы. Это различие сохраняется и в культуре при одинаковых условиях выращивания. Экспериментальное вытаптывание растений различного происхождения (для этого на них многократно опускали металлические гири подходящего размера так, чтобы производимое ими давление на единицу поверхности примерно соответствовало давлению, оказываемому взрослым человеком) показало, что стелющиеся цветоносы по приспособленности (измеренной как сухой вес репродуктивных органов) статистически достоверно слабее реагируют на искусственное вытаптывание, чем прямостоячие. В контроле (без вытаптывания) приспособленность распростертых и прямостоячих форм оказалась почти одинаковой. В эксперименте с реципрокными пересадками поменяли местами популяции одного вида из контрастных местообитаний. Так, например, прибрежные популяции калифорнийского Achillea lanulosa были пересажены в горы, а высокогорные — на побережье. В обоих случаях было обнаружено, что приспособленность популяций в чуждом местообитании ниже, чем в естественном. Оба эксперимента позволяют сделать вывод, что наблюдаемые в природе различия между популяциями, обитающими в разных экологических условиях, являются результатом естественного отбора и могут интерпретироваться как адаптивные различия. Реципрокное пересаживание имеет то преимущество перед экспериментальным анализом только одного фактора окружающей среды, которое позволяет измерить приспособленность ко всей среде в целом, не устанавливая часто трудно идентифицируемого фактора среды, предположительно наиболее важного для изменчивости.
Совершенно не ясно, можно ли каждый фенотипический признак организма интерпретировать как адаптацию. Существует альтернативная вероятность, что возникающие благодаря мутациям признаки селективно нейтральны и сохраняются благодаря особым генетическим процессам (например, при плейотропии, генетическом сцеплении). Можно ожидать, что мутации, приводящие к селективно нейтральному признаку (поскольку они не влияют на приспособленность), происходят сравнительно часто и медленно отсеиваются естественным отбором. Известное постоянство признаков указывало бы, таким образом, на то, что ограниченная изменчивость признака — результат естественного отбора. Соответственно нельзя с уверенностью принять, что каждое свойство организма является совершенной адаптацией. Следует думать, что структура или свойство чаще всего имеет многообразные связи с окружающей средой и, соответственно, может быть понята как результат действовавшего в различных направлениях естественного отбора, т. е. как некий компромисс. Одно какое-либо свойство, кодируемое плейотропно действующим геном или комплексом тесно сцепленных генов, может поддерживаться естественным отбором, тогда как другое свойство, кодируемое тем же геном или тем же комплексом, — нет. Наличие последнего тогда не адаптивно. Наконец, может быть и так, что какое-то свойство является приспособлением к уже не существующим условиям среды, а времени для его эволюционного изменения прошло слишком мало. Так, например, очень крупные и жесткие плоды многочисленных южноамериканских видов деревьев (например, Crescentia alata, Bignoniaceae; Scheelea rostrata, Arecaceae) интерпретируют как приспособление к крупным травоядным животным, вымершим примерно 10000 лет назад.
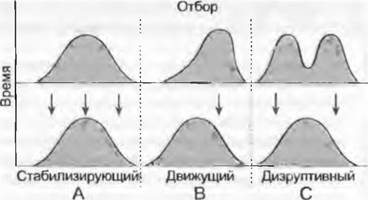
В зависимости от эффекта эволюционного изменения признака различают три формы естественного отбора: движущий, дизруптивный и стабилизирующий (рис. 10.28). В случае движущего отбора в популяции с нормальным распределением генетической изменчивости наиболее приспособленными оказываются одни из крайних вариантов. Это приводит в последовательных поколениях к направленному смещению изменчивости вариационной кривой в сторону этой краевой области. При дизруптивном отборе в популяции генотипы обеих краевых зон вариационной кривой имеют более высокую приспособленность, чем генотипы индивидов средней части спектра изменчивости. В результате начинается разделение популяции на две. В случае стабилизирующего отбора, наоборот, генотипы в области средних популяционных показателей имеют наибольшую приспособленность, а генотипы в обеих краевых зонах — низкую. Такое распределение уровня приспособленности индивидуумов не изменяет генетическую изменчивость в ряду последовательных поколений. На примере стабилизирующего отбора очевидно, что эволюционным результатом естественного отбора не всегда должно быть изменение частот аллелей.
Рис. 10.28. Стабилизирующий, движущий и дизруптивный отбор. Размах вариации (абсцисса) исходных популяций (внизу) обусловлен частотой индивидуумов с различной наследственностью: он либо сохраняется (А), либо сдвигается (В), либо разделяется на две части (С) благодаря разным формам отбора (стрелки)
Внутривидовая генетическая изменчивость коррелирует не только с факторами окружающей среды. Анализ генетической изменчивости, в особенности на молекулярном уровне (например, изоферменты), показал, что данный тип генетической изменчивости частично коррелируют и обусловлен теми свойствами, которые в совокупности составляют рекомбинационную систему вида. Здесь прежде всего интересно сравнение внутрипопуляционной и межпопуляционной изменчивости. Система опыления в целом действует так, что самоопыляющимся видам в основном присуща сравнительно малая внутрипопуляционная, сравнительно большая межпопуляционная изменчивость. У перекрестноопыляющихся видов соотношения обратные — внутри популяций изменчивость выше, чем между популяциями. Объясняются эти примеры, с одной стороны, генетическим эффектом постоянного самооплодотворения, с другой стороны, различной интенсивностью генетического обмена (потока генов) между популяциями у само- и перекрестноопыляющихся видов. Самооплодотворение ведет к утрате генетической изменчивости внутри популяции и одновременно способствует изоляции соседних популяций. Влияние потока генов на структуру внутривидовой генетической изменчивости ясно также при сравнении разных механизмов переноса пыльцы и диаспор. Ветроопыляемым, распространяемым ветром видам свойственны сравнительно большая внутрипопуляционная и сравнительно меньшая межпопуляционная изменчивость, чем видам, опыляемым и распространяемым животными, и видам без особых приспособлений для распространения. Создается впечатление, что дистанции потока генов и соответственно генетический обмен между популяциями у ветроопыляемых и распространяемых ветром видов в среднем больше, чем у видов, опыляемых и распространяемых животными. На проявление изменчивости влияет и жизненная форма: однолетние виды демонстрируют меньшую изменчивость внутри популяций и большую между популяциями, нежели короткоживущие травянистые многолетники, а последние, в свою очередь, — меньшую внутрипопуляционную и большую межпопуляционную изменчивость, чем долгоживущие древесные многолетние виды. Возможное объяснение этих примеров лежит, с одной стороны, в известной корреляции самоопыления и малого потока генов с короткой продолжительностью жизни, а перекрестного опыления и более сильного потока генов с большой продолжительностью жизни, а с другой стороны, в том, что со временем популяции долгоживущих видов с большей вероятностью обогащаются генетически благодаря притоку генов из других популяций, чем популяции короткоживущих видов. Наконец, у видов с бесполым или частично бесполым размножением генетическая изменчивость существенно меньше, чем у видов только с половым размножением.
Бокс 10.2. Популяционная гонетика
Изменение частот аллей в последовательных поколениях количественно исследуют в популяционной генетике. Популяционная генетика пытается таким образом ответить на вопрос: какими станут частоты генотипов или соответственно аллелей в каком-либо поколении, если известны их частоты в предыдущем поколении? Частоту аллелей можно выяснить из рассчитываемой частоты генотипов. В случае единственного генетического локуса с аллелями А и а имеется три генотипа — АА, Аа и аа с частотами Р, Q и R. Частоту р аллеля А рассчитывают делением суммы удвоенной частоты гомозиготных генотипов АА и частоты гетерозиготных генотипов Аа на удвоенное число индивидуумов в популяции:
![]()
Следует использовать удвоенную частоту генотипа АА и удвоенное число индивидуумов, потому что каждый диплоидный индивидуум содержит два аллеля, так что число аллелей в популяции вдвое больше числа индивидуумов. Так как частоты Р, Q и R трех генотипов в сумме дают частоту всех индивидуумов в популяции и равны 1, данную выше формулу можно упростить: р = Р + Q/2. Частоту q аллеля а рассчитывают аналогичным образом по формуле q = R + Q/2. Поскольку частоты обоих аллелей в сумме равны 1 (р + q = 1), q можно вычислять так: q = 1 - р. Напротив, частоту генотипов по частотам аллелей можно вычислить только при определенных условиях. Рассчитать частоту генотипов в следующем поколении по частотам аллелей в предыдущем поколении возможно, если:
✵ приспособленность трех генотипов АА, Аа и аа (как в примере) одинакова,
✵ генотипы скрещиваются случайно,
✵ не появляются новые аллели (из-за мутаций или потока генов),
✵ популяция достаточно многочисленна, чтобы исключить случайные флуктуации частот аллелей.
В этом случае частота АА равна р2, частота Аа равна 2рq и частота аа равна q2. Эта основополагающая для популяционной генетики связь между частотой аллеля и частотой генотипа известна как закон Харди — Вайнберга. Он означает, что при постоянных условиях в череде последовательных поколений не происходит сдвига частот аллелей и генотипов и, соответственно, нет эволюции. Однако можно проверить, находится ли популяция в равновесии или нет. Из частоты имеющихся в популяции генотипов АА, Аа и аа можно рассчитать частоты р аллеля A и q аллеля а. Если популяция находится в равновесии, то частоты генотипов АА, Аа и аа должны оказаться р2, 2рq и q2. Если наблюдаемые частоты генотипов отклоняются от ожидаемых согласно закону Харди—Вайнберга, то популяция не находится в равновесии. Причиной этого могла бы быть неодинаковая приспособленность трех генотипов. Можно рассчитать, как неодинаковая приспособленность влияет на частоту аллелей в последующих поколениях. Для простоты можно принять, что приспособленности АA и Аа одинаковы и равны I, но приспособленность аа меньше: I - s. Буквой s обозначают коэффициент отбора, который равен единице минус приспособленность, причем в простейшем случае приспособленность можно рассчитать, как отношение уровня выживания данного генотипа к уровню выживания наиболее приспособленного генотипа. В нашем примере генотипы АА и Аа имеют приспособленность I, а уровень выживания генотипа аа мог быть 90 % от таковой АА и Аа. Тогда его приспособленность была бы 0,9, а коэффициент отбора s = 0,1. В исходной популяции частоты гентипов составляют АА = р2, Аа = 2рq и аа = q2(1 - s). Общая величина популяции: р2 + 2рq + q2(I - s). Если в этой формуле заменить р на 1-q (р + q =1, откуда р = 1 - q), то выражение для общей величины популяции можно упростить: 1 - sq2. Тогда частоты генотипов в популяции будут:
![]()
Частота р1, изменившаяся под действием отбора, будет:
![]()
(Частота АА + половина частоты Аа как доли общей популяции.) Изменение частоты в ходе отбора:
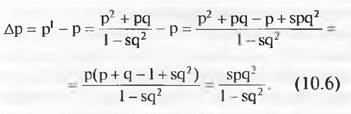
Если первоначально р = q = 0,5 и s = 0,1, то

Таким образом, частота р, равна 0,5128, а частота q1 равна 0,4872. Эта формула дает возможность рассчитать и предсказать изменение частоты аллеля в последовательных поколениях при случайном скрещивании и разной приспособленности генотипов. Если принять, что коэффициент отбора гомозиготного рецессивного генотипа аа = 1, т. е. индивидуумы этого генотипа никогда не достигают репродуктивного состояния, то по исходной частоте q0 аллеля а можно рассчитать частоту а qn через n поколений:
![]()
При исходной частоте qo = 0,5 частота qn аллеля а после, к примеру, 10 поколений (n = 10) постоянной элиминации станет q10 = 0,083. Отсюда ясно, что даже очень низкая приспособленность гомозиготного фенотипа не может привести к полной элиминации обусловливающего его аллеля, так как данный аллель содержится в гетерозиготных генотипах. Это подтверждает большое эволюционное значение диплоидности и связанной с ней гетерозиготности большого числа организмов. Сложнее сделать подобные расчеты в тех случаях, когда генотипы различаются не уровнем выживания, а репродуктивным успехом, когда скрещивания в популяции неслучайны, когда имеет место поток генов между популяциями или, когда приспособленность генотипов изменяется в ряду поколений, например, в зависимости от их частоты. Еще более усложняется количественный анализ, когда наблюдают отбор не по одному генетическому локусу, а по двум или более локусам, взаимно влияющим друг на друга. В то же время это более соответствует реальной ситуации, так как, пожалуй, лишь немногие фенотипические особенности кодируются только одним генетическим локусом.
10.2.2. Дрейф генов
Другой важный для генетической структуры видов фактор — случайность. Случайные сдвиги частот аллелей в следующих друг за другом поколениях называют дрейфом генов. К таким событиям может, в частности, привести резкое сокращение размера популяции, причем с уменьшением популяции возрастает вероятность случайного сдвига частот аллелей. В результате дрейфа генов может произойти утрата одного из аллелей, и благодаря этому, по-другому аллелю одного генетического локуса все индивидуумы оказываются гомозиготными (фиксация). Правда, генетический эффект уменьшения численности популяции становится заметным только тогда, когда ее величина становится меньше 10 индивидуумов и в течение нескольких поколений остается низкой, вследствие чего в основном происходит инбридинг, а один из аллелей локуса был редким в исходной популяции. Величина популяции может быть малой, например, из-за того, что немногие индивидуумы или даже всего один индивидуум (гермафродитный и самооплодотворяющийся) оказались на острове в результате дальнего заноса или случайного заноса человеком и основали новую популяцию (эффект основателя, англ. founder effect). Резко сократить численность популяции также могут и драматические изменения среды (эффект бутылочного горлышка, англ. bottleneck).
Пример эффекта основателя — занесенный человеком из Северной Америки в Австралию злак Echinochloa microstachya. В то время как в Северной Америке каждая популяция этого вида отличается от других характерной для нее комбинацией аллелей, 18 из 20 изученных в Австралии популяций оказались генетически идентичными. Однако подобное единообразие встречается не всегда. Так, занесенная из Европы в Северную Америку метлица (Apera spicaventí) демонстрирует на обоих континентах сходный размах генетической изменчивости.
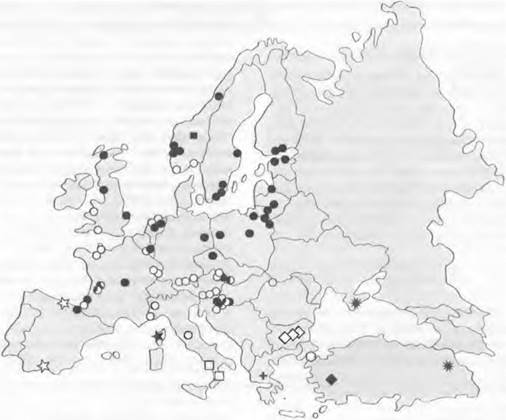
Постоянное изменение высоты ледяного покрова в четвертичном периоде и связанные с этим климатические изменения сильно влияли на величину популяций всех живших тогда организмов. Этим можно объяснить, что в областях- убежищах (рефугиумах) многочисленные виды деревьев характеризуются сравнительно высокой генетической изменчивостью, а в колонизированных ими в конце ледникового периода областях на Севере популяции генетически обеднены (рис. 10.29). Такой тип генетической изменчивости мог возникнуть благодаря тому, что повторное заселение подвергшейся оледенению территории не шло путем плавного расширения ареала прежде отступивших популяций. В уже заново заселенных областях всегда происходили колебания численности, обусловленные быстрыми изменениями климата, либо единичные или немногие индивидуумы могли натурализоваться далеко впереди от наступающей популяции.
Рис. 10.29. Географическое распространение 11 различных типов пластид Alnus glutinosa. В ледниковых рефугиумах в Южной Европе находят большее генетическое разнообразие, чем во вновь заселенных оттуда областях. Кружки с черными и белыми сегментами указывают на гетерогенность популяций по типам пластид
Наблюдения, позволяющие объяснить характер генетической изменчивости определенными особенностями растений (например, системой оплодотворения) или случайными событиями, поднимают вопрос о роли естественного отбора. Более общий вопрос — в какой мере генетическая изменчивость нейтральна, т. е. в какой мере различные генотипы имеют одинаковую приспособленность. Относительно фенотипических признаков можно в целом принять, что различные генотипы нередко имеют и разную приспособленность. Поэтому характер генетической изменчивости по фенотипическим признакам часто является результатом естественного отбора. Относительно молекулярных признаков1 было постулировано, прежде всего М.Кимурой, что большинство имеющихся в популяции генотипов обладают одинаковой приспособленностью и потому селективно нейтральны. В то же время нет никакого сомнения в том, что эффект большинства новых мутаций (см. рис. 10.29) отрицательный, так как эти мутации элиминируются отбором. Кимура предположил: генетическая изменчивость на молекулярном уровне — скорее результат дрейфа генов, нежели естественного отбора.
1 Исключение молекулярных признаков из числа фенотипических — ошибка. — Примеч. ред.
Эта теория нейтральной эволюции основывается на слишком высоких для результатов естественного отбора темпах эволюции протеинов, слишком большом многообразии протеинов внутри видов, на известном постоянстве темпов эволюции протеинов в разных эволюционных линиях и на более высоких темпах эволюции нефункциональных частей ферментов в сравнении с функциональными. Аргументация, первоначально базировавшаяся на анализе протеиновых последовательностей, позднее была изменена благодаря несколько иным результатам анализа нуклеотидных последовательностей ДНК.
Ответ на вопрос, что влияет сильнее на молекулярную эволюцию — дрейф генов или естественный отбор, зависит в значительной мере от представлений о размере популяций, темпах мутационного процесса и коэффициентах отбора. Поскольку эти величины обычно не удается определить количественно и им приходится давать только приблизительную оценку, то в зависимости от принятых оценок определенные варианты генетической изменчивости можно объяснить в соответствии с нейтралистской и селекционистской моделями эволюции. Соответственно нельзя окончательно ответить на вопрос об относительной роли дрейфа и отбора. Достоверно лишь то, что, к примеру, скрытые мутации или некоторые нетранскрибируемые последовательности ДНК эволюируют нейтрально и что на молекулярную эволюцию, следовательно, влияют как дрейф, так и отбор. Но и в фенотипической эволюции может проявляться дрейф генов.