БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
10. ЭВОЛЮЦИЯ
10.3. Видообразование
10.3.1. Определения вида
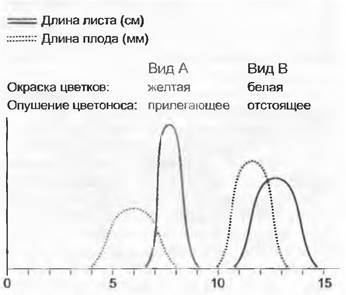
Обсуждение генетической изменчивости внутри вида и механизмов видообразования с очевидностью требует определения вида. Дать непротиворечивое определение вида чрезвычайно трудно. Несомненно, в практической систематике большинство видов описано на основе наблюдаемой морфологической изменчивости и, таким образом, использована морфологическая (таксономическая или фенотипическая) концепция вида. При этом систематик ищет скоррелированную прерывистость в изменении разных фенотипических, главным образом морфологических, признаков (рис. 10.30). Эту прерывистость рассматривают как границу между видами. Изменчивость внутри видов непрерывна, а изменчивость между видами прерывиста. Однако следует иметь в виду, что в особых случаях и внутривидовая изменчивость бывает прерывистой в форме полиморфизма (например, гетеростилия у Prímula, двудомность; см. 10.1.3). В подобных ситуациях для разграничения видов требуются дополнительные критерии. Именно по этой причине в понимании морфологического вида сильна субъективная компонента, ибо невозможно установить объективные критерии величины фенотипической прерывистости, необходимой для разграничения видов.
Видовые границы могут быть по-разному выражены в разных пространственных масштабах. Разграничение видов в узкоограниченной области чаще всего вызывает немного проблем, но они существенно возрастают при рассмотрении всей области распространения вида (см. рис. 10.30).
Рис. 10.30. Схематическое изображение дискретной изменчивости между видами по количественным и качественным признакам. Виды А и В различаются по окраске цветков, опушению цветоносов, а также по длине листьев и плодов
В то время как морфологическое определение вида в принципе не связано с представлениями об эволюционном процессе, в других определениях вида пытаются учесть процесс его эволюционного образования. Сформулированная Э. Майром биологическая концепция вида определяет виды как группы свободно скрещивающихся природных популяций, которые репродуктивно изолированы от других групп популяций (т. е. от других видов). Репродуктивная изоляция имеет генетическую основу. Это определение подразумевает, что виды распознаваемы потому, что они не гибридизируют с другими видами. Гибридизация как критерий распознавания вида уже давно и неоднократно обсуждалась, и использовалась Э. Майром.
В биологической концепции вида подчеркивается именно репродуктивная изоляция одного вида от другого. Очень похожая концепция взаимоузнавания конспецифичных партнеров (англ. recognition species concept), напротив, подчеркивает взаимное узнавание партнеров по скрещиванию внутри вида. Экологическая концепция вида определяет вид как группу популяций, которые занимают одинаковые экологические ниши. Принимается, что интеграция видов возникает на основе сходной экологии принадлежащих к ним индивидуумов и популяций, и соответственно, сходного направления отбора. Генетическая концепция вида пытается объяснить существование видов тем, что они могут изменяться только в определенных границах, определяемых их собственной генетической структурой.
В то время как вышеназванные концепции вида применимы к ныне существующим организмам, другие концепции пытаются определить вид в его историческом существовании. Эволюционная концепция вида Дж. Симпсона определяет вид как линию развития (т. е. последовательность возникающих друг от друга популяций), которая эволюирует независимо от других подобных линий развития и имеет собственную эволюционную роль и тенденцию развития.
И наконец, в различных вариантах филогенетической (кладистической) концепции вида в один вид объединяют всех произошедших от одного предка членов одной (монофилетической) эволюционной линии развития — от ее возникновения (образования вида) до конца (т. е. образования следующих видов). Таким образом, здесь в основу для распознавания видов положено общее происхождение от одного предка и неявно предполагается, что виды возможно различать только благодаря их собственным признакам (аутапоморфиям).
Очевидно, что различные концепции вида были сформулированы на основе разных, но отнюдь не взаимоисключающих теоретических положений. Независимо от их теоретической ценности они в очень разной степени годятся для практического использования. Выделенные согласно разным концепциям виды наверняка нередко широко перекрываются. Можно заключить, что морфологические виды существуют потому, что принадлежащие к ним индивидуумы скрещиваются между собой, но репродуктивно изолированы от других видов, что они образовались при одинаковом давлении отбора, являются результатом независимой эволюции и возникли от одного общего предка. Поскольку нет общепринятого критерия видового ранга, морфологическую концепцию вида иногда рассматривают как ненаучную.
Прежде чем давать определение вида, необходимо ответить на вопрос о правомерности объединения индивидуумов и популяций в виды как реально существующие в природе основные единицы биологической изменчивости. В зависимости от той или иной точки зрения на этот вопрос можно ответить по-разному. Так как основной единицей эволюционного процесса является популяция как группа живущих на определенной территории и скрещивающихся между собой индивидуумов — ибо именно в популяциях имеет место сдвиг частот аллелей в череде поколений, — можно сделать вывод, что сам ход эволюционного процесса не позволяет признать виды за естественные целостности, и виды, так же, как и таксономические единицы более высокого ранга (роды, семейства и т. п.), — категории искусственные. К этому выводу пришел уже Ч. Дарвин. С другой стороны, правда и то, что существование скрещивающихся между собой популяций и их репродуктивная изоляция от других популяций — биологическая реальность, что несколько популяций подвержены сходному давлению отбора благодаря сходной экологии и что несколько популяций, обладающих общими, только им свойственными признаками, произошли от общего предка. При таком подходе объединение индивидуумов и популяций в виды отражает биологическую реальность.
Подводя итог, можно сказать, что систематическая категория «вид» независимо от трудности или даже невозможности ее удовлетворительного определения является до сих пор незаменимым основным понятием для оперирования с научными наблюдениями. Так как на практике большинство видов можно опознать по морфологическим признакам и очень многие «морфологические» виды соответствуют также и критериям других концепций вица, здесь следует понимать виды именно как морфологиче
ские, которые обычно изолированы друг от друга генетическими механизмами. При этом нельзя отрицать, что морфологическая концепция вида в известной мере субъективна и теоретически недостаточна. Особая проблема распознавания видов возникает у агамоспермных форм (см. 10.1.3.3).
Таким образом, дискуссия о процессе видообразования должна объяснить, как в эволюции возникает, с одной стороны, морфологическое разнообразие, а с другой — репродуктивная изоляция.
10.3.2. Дивергентное видообразование
10.3.2.1. Аллопатрическое видообразование
Внутривидовая изменчивость возникает преимущественно благодаря мутациям, естественному отбору и дрейфу генов. Если генетический обмен между популяциями или популяционными группами одного вида прерывается, например, из-за их пространственного разделения или вымирания промежуточных популяций, то процесс внутривидовой дифференциации в генетически изолированных частях вида может продолжаться благодаря различным мутациям, приспособлению к разным условиям среды в ходе естественного отбора и/или случайной фиксации разных аллелей и привести к возникновению новых видов. Эволюционный процесс разделения одного вида на два (или несколько) новых называется видообразованием (англ. spéciation). Простейшая возможность прекращения потока генов между популяциями одного вида — пространственное разделение. Соответственно этот механизм видообразования называют также географическим, или аллопатрическим, видообразованием. В данном случае усиливающаяся дивергенция изолированных популяций происходит постепенно, и принято считать, что дивергенция обусловлена преимущественно естественным отбором.
Пространственное разделение и связанная с ним генетическая изоляция популяций одного вида могут, к примеру, возникнуть вследствие разделения массивов суши при движении континентов, возникновения горных систем, фрагментации ареалов (при четвертичных оледенениях) или разделения массивов суши водными преградами из-за потепления климата в послеледниковье (например, Британские острова/Европейский континент). Геологические и климатические изменения в истории Земли бессчетное число раз вызывали расчленение прежде непрерывных ареалов и создавали тем самым предпосылки для видообразования.
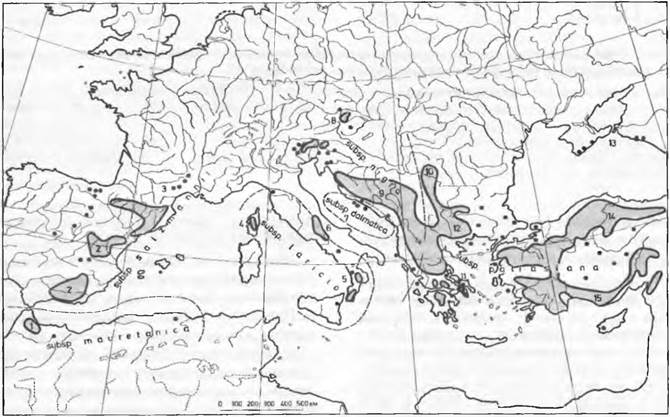
Ввиду почти полной невозможности непосредственно наблюдать процесс видообразования в природе важнейшим указанием на большое значение аллопатрического видообразования оказывается географическая специфика внутривидовой изменчивости, соответственно — географическое распространение близкородственных видов. Так, например, у обитающей в Средиземноморье сосны черной (Pinus nigra) обнаружены многочисленные географически изолированные подвиды (рис. 10.31), а внутри подвидов отмечена географическая дифференциация. Дифференциацию внутри подвидов и сами подвиды можно рассматривать как разные стадии непрерывного процесса аллопатрического видообразования.
Части вида называют подвидами в том случае, если большинство индивидов можно отнести к тому или иному подвиду (см. рис. 10.31), но существуют и редкие промежуточные формы. Таким образом, изменчивость между подвидами не полностью прерывиста. Кроме того, важно, что подвиды либо обитают в разных областях, либо различаются экологией.
Рис. 10.31. Географическая дифференциация круга горных средиземноморских форм сосны черной (Pinus nigra). Подвиды поименованы, соподчиненные локальные расы обозначены цифрами
В группе видов Gilia (Polemoniaceae), распространенных на юго-западе Северной Америки, находят географически обособленные (С. leptantha) или граничащие друг с другом (G. latiflora) близкородственные виды с частично перекрывающимися ареалами (С. tenuiflora, G. latiflora, C. leptantha), а также менее близкородственные аллопатрические (С. ochroleuca,G. mexicana) либо симпатрические виды (G. ochroleuca, G. tenuiflora, G. leptantha, G. latiflora) (рис. 10.32).
Рис. 10.32. Видообразование и географическое распространение в одной из групп видов рода Gilia (Polemoniaceae) на юго-западе Северной Америки. Существуют географически граничащие друг с другом (А — G. latiflora) или изолированные (В — G. lерtantha) подвиды внутри видов, близкородственные и частично перекрывающиеся виды (С — G. lentiflora, G. latiflora, G. leptantha), менее близкородственные либо аллопатрические (D — G. ochmleuca, G. mexicana) либо симпатрические виды (Е — G. ochroleuca, G. tenuiflora, G. leptantha, G. latiflora). Этот пример географического распространения и различной степени родства интерпретируют как видообразование на различных стадиях
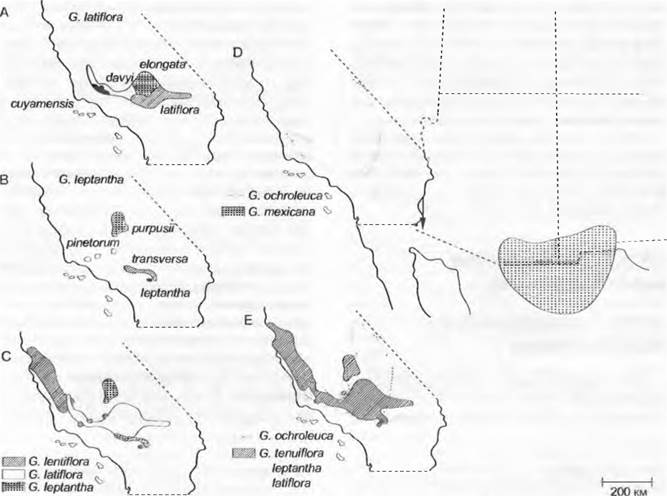
Такой характер географического распространения и разной степени родства интерпретируют как разные стадии видообразования. Подобные типы распространения и родственных связей можно найти почти повсеместно. Gilia ochroleuca не способна скрещиваться с встречающимися в той же области G. tenuiflora, G. leptantha, G. latiflora (см. рис. 10.32). Но последние три вида с частично перекрывающимися ареалами время от времени скрещиваются между собой. Эти наблюдения показывают, что аллопатрически возникающие виды могут унаследовать механизмы репродуктивной изоляции, которые позволяют совместно обитать близкородственным видам.
Правда, следует задаться вопросом: действительно ли такое распространение родственных видов отражает процесс аллопатрического видообразования и им обусловлено, разве не могло оно возникнуть вследствие совсем других причин уже после образования видов?
Постепенность аллопатрического видообразования проявляется, например, в более или менее непрерывном увеличении генетического расстояния, выявляемого при сравнении изоферментов, между популяциями одного подвида, подвидами одного вида или близкородственными видами, более далекими видами и т.д.
10.3.2.2. Репродуктивная изоляция
Возможные механизмы репродуктивной изоляции, обусловленные своеобразием растений, можно сгруппировать в зависимости от момента их действия. При этом самые важные моменты — это опыление и образование зиготы. Уже попадание на рыльце индивида большого количества пыльцы другого вида (рис. 10.32) может препятствовать его репродуктивному успеху. Образование зиготы при оплодотворении яйцеклетки спермием чужого вида ведет к бесплодию и тем самым снижает приспособленность индивидуума. Действенные механизмы репродуктивной изоляции, включающиеся перед опылением (англ. pre-mating) и оплодотворением (презиготические), являются важной предпосылкой аллопатрического видообразования наряду с уже обсуждавшейся пространственной изоляцией.
• Экологическая изоляция. Почти полностью перекрывающиеся ареалами виды гравилат городской (Geum urbanum) и гравилат речной (С. rivale) встречаются соответственно во влажных смешанных лесах, кустарниковых зарослях, по опушкам и в сырых местообитаниях различного рода. Если их местообитания оказываются рядом, то виды часто гибридизируют.
• Временная изоляция. Время цветения близкородственных и встречающихся вместе видов может быть разным. Пример суточного различия времени цветения — цветущий весной Lactuca graminifolia и цветущий летом L. canadensis на юго-востоке США. Пример суточного различия цветения — Silene latifolia (цветки раскрываются ночью) и S. dioica (цветки раскрываются днем); они также изолированы благодаря разным опылителям.
• Изоляция экологией цветения. Гибридизации близкородственных видов могут препятствовать специфические опылители. Яркий пример — орхидеи рода Ophrys, строение и аромат цветков разных видов которых имитируют самок разных перепончатокрылых и их феромоны и потому посещаются самцами разных видов. Близкородственные губастик красный (Mimulus сагdinalis) и губастик Льюиса (М. lewisit) (Scrophulariасеае) в Калифорнии опыляют соответственно колибри и шмели. С различием опылителей обычно связаны также и морфологические различия цветков, которые более или менее исключают опыление неспецифичным опылителем. Данное явление называют также механической изоляцией.
Другой случай изоляции экологией цветения — связанная с видообразованием (или внутривидовой дифференциацией) замена системы самонесовместимости и перекрестного опыления на самосовместимость и самооплодотворение. Это особенно четко выявлено на примере эволюции от связанной с тристилией самонесовместимости к почти гомостилии и самосовместимости в роде Еichhornia (Роntederiасеае). Здесь репродуктивную изоляцию между видами обеспечивают самоопыление и самооплодотворение.
Успешному оплодотворению и образованию зиготы после опыления (англ. роst-mading) может препятствовать так называемая гибридная несовместимость.
• Гибридная несовместимость. Ее обеспечивают механизмы, препятствующие прорастанию пыльцы, росту пыльцевой трубки через столбик, выходу ядра спермия из пыльцевой трубки или оплодотворению вторичного ядра зародышевого мешка, необходимого для образования эндосперма у цветковых растений.
Другой аспект роста пыльцевых трубок состоит в том, что при попадании на рыльце собственной и чужой пыльцы пыльцевые трубки собственной пыльцы обычно растут быстрее, чем пыльцевые трубки чужеродной пыльцы. В зависимости от количественных соотношений между собственной и чужой пыльцой в той или мере снижается успех оплодотворения пыльцой чужого вида.
После образования зиготы включаются постзиготические механизмы.
• Сниженная жизнеспособность гибридов F1. Пониженная жизнеспособность гибридных индивидуумов может проявляться на разных стадиях — от первого деления зиготы до цветущего и плодоносящего растения.
• Стерильность гибридов. Фертильность гибридов может быть снижена благодаря генным, хромосомным или цитоплазматическим различиям между родительскими видами. Различия по одному или многим генам могут привести при их комбинировании у гибридов к снижению фертильности.
Пример хромосомной мутации (инверсия) как причины сниженной фертильности гибридов между индивидуумами с мутацией и без нее представлен на рис. 10.8. Удалось показать, что сниженная фертильность гибридов между двумя видами подсолнечника (Helianthus annuus и Н. argophyllus) обусловлена различием родительских видов по двум реципрокным транслокациям. Пониженную фертильность гибридов могут обусловить также различия в числе хромосом, возникшие в результате анеуплоидии, дисплоидии или полиплоидии. Понятно, что возникновение хромосомных различий внутри вида представляет исходный момент для видообразования. При сравнении близкородственных видов во многих случаях четко выявлены хромосомные мутации. Проявляющиеся в стерильности гибридов генные и хромосомные различия чаще всего вызывают нарушения в образовании бивалентов в мейозе у гибридных индивидуумов.
• Гибридный распад. Снижение жизнеспособности или фертильности проявляется не в первом гибридном поколении, а в последующих.
Концепция аллопатрического видообразования предполагает, что названные механизмы изоляции являются побочным продуктом возрастающей генетической дивергенции образующихся видов.
Разную степень репродуктивной изоляции использовали в экспериментальной систематике в трудоемких опытах по скрещиванию для определения степени родства между видами. При этом, правда, стало очевидно, что эволюционная дивергенция по морфологическим признакам и репродуктивная изоляция вовсе не всегда тесно сцеплены. Известны примеры, когда морфологически очень разные виды могут легко скрещиваться и давать фертильные гибриды, а также примеры, где едва различимые формы репродуктивно изолированы друг от друга.
10.3.2.3. Перипатрическое, парапатрическое, симпатрическое видообразование и эффект основателя
Серьезная критика модели аллопатрического видообразования основана на представлении о том, что весьма трудно идентифицировать факторы, ответственные за общую эволюцию множества популяций, населяющих часть области распространения вида. Эта общность эволюции была бы возможной при условии потока генов между популяциями и одинакового направления отбора во всей данной области. На это следует возразить, поскольку дистанции потока генов в природе чаще всего малы (см. 10.1.3.4), а условия окружающей среды обычно варьируют на небольших расстояниях, популяции приспособлены к локальным условиям. На основе данного соображения предложены модели видообразования, в которых за исходную точку берут малые, но пространственно изолированные популяции вида. Один из примеров — перипатрическое видообразование (по-английски иногда его называют quantum spéciation), при котором обитающие на краю ареала популяции из-за отклоняющихся условий среды оказываются изолированными и могут дать начало новому виду. Важнейшие аргументы в пользу этой модели видообразования — опять же специфика географического распространения внутривидовых вариаций или близкородственных видов. В принципе похожая модель видообразования, названная эффектом основателя (англ. founder effect spéciation), не содержит указаний на то, как малая популяция стала изолированной. Здесь, в частности, возможно, что диаспоры вида попали в географически изолированную область, например, на остров. Для обеих моделей общим является внимание к эволюционным процессам при возникновении и после возникновения малых популяций: к уменьшению генетической изменчивости при образовании малой популяции (дрейф генов; см. 10.2.2), а также к быстрой эволюции и возникновению новых особенностей благодаря инбридингу после образования популяции. Это подразумевает, что при видообразовании в малых популяциях случайные процессы имеют большее значение, чем естественный отбор.
Примеры возможного перипатрического видообразования — пары видов Lasthenia т'тог / L. maritima (Asteraceae) и Salix alaxensis / S. sili- cicola (Salicaceae). Lasthenia minor населяет материковую часть Калифорнии, тогда как L. Maritima чаше всего растет на скалистых островках с птичьими базарами у калифорнийского побережья. Предположили, что L. maritimaвозникла из краевых популяций L. minor, изолированных от материка в конце четвертичного периода в результате подъема уровня моря. В то время как арктический вид Salix alaxensis широко распространен в западной Канаде и на Аляске, A. silicicola имеет узкий ареал в дюнах, образовавшихся после ледникового периода по южному краю озера Атабаска в Канаде. Ареалы обоих видов в настоящее время разделяет примерно 300 км. Возможно, S. silicicola возникла после отступления ледника в конце четвертичного периода из сохранившихся у озера Атабаска популяций мигрировавшей на северо-запад S. alaxensis. Как Z. maritima. так и 5. silicicola отличаются генетически от их предполагаемых предков L. minor и S. alaxensis прежде всего утратой генетической изменчивости.
Примеры эффекта основателя виды Cirsium canescens /С. pitcheri (Asteraceae) и Sanícula (Apiaceae) на Гавайских островах. Подобно Lasthenia и Salix распространенный по берегам Великих озер Северной Америки С. pitcheri проявляет лишь малую часть генетической изменчивости, характерной для широкораспространенного в Небраске С. canescens.Предположили, что предок С. pitcheri мигрировал во время последнего оледенения вдоль края ледника к Великим озерам и таким образом оказался изолированным от С. canescens. Sanícula на Гавайях — один из многих примеров видообразования после однократного заселения островов немногими диаспорами. Четыре вида Sanícula, встречающиеся на трех островах Гавайского архипелага, близкородственны калифорнийским видам этого рода. Обнаружение сниженной по сравнению с материковыми видами генетической изменчивости как у всей группы четырех гавайских видов, так и у каждого из них. а также интерпретация кладистических расстояний последовательностей ДНК как молекулярных часов позволяют предположить, что около 8,9 млн лет назад архипелага достигли всего один раз немногие диаспоры (или даже единственная) и что дальнейшее видообразование происходило на архипелаге по модели эффекта основателя примерно 0,9 млн лет назад. Видообразование на архипелаге — пример адаптивной радиации, при которой за колонизацией нового жизненного пространства чаще всего быстро следует эволюционная диверсификация.
Если бы видообразование происходило только в малых популяциях, то географическое распространение, например, подвидов Pinus nigra (см. рис. 10.31), можно было
бы объяснить и тем, что новые таксоны, возникнув в малых популяциях путем пе- рипатрического образования подвидов или видов, или благодаря эффекту основателя, увеличили свои ареалы.
Бесспорно, географически, а тем самым и генетически изолированные части вида могут дать начало новым видам. Однако спорным остается вопрос о том, могут ли стать новыми видами непосредственно граничащие друг с другом (парапатрические) или распространенные на одной и той же территории (симпатрические) популяции одного вида. Поскольку прекращение потока генов — важная предпосылка для дивергентной эволюции, в таких случаях был бы необходим механизм репродуктивной изоляции. Этот механизм может возникнуть, если гибриды между соседними, но адаптированными к разным условиям среды популяциями обладают резко пониженной приспособленностью.
Рис. 10.33. Симпатрическое видообразование. В эксперименте с кукурузой (Zea mays) интенсивный отбор против гибридизации между двумя сортами (White flint, Yellow sweet) с одинаковыми сроками цветения после 6 поколений приводит к отчетливому расхождению по срокам цветения
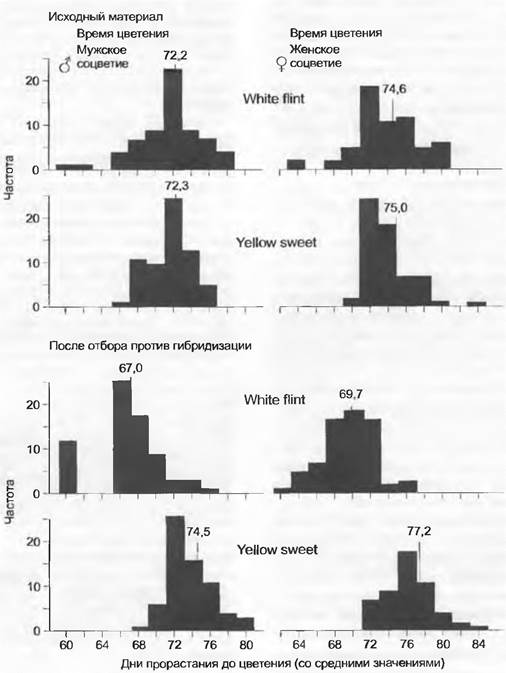
Возникновение репродуктивной изоляции в симпатрических популяциях благодаря сдвигу во времени цветения было показано в селекционных экспериментах с двумя сортами кукурузы (Zea mays) (рис. 10.33). Исходным материалом были сорта кукурузы с желтыми (Yellow sweet) и белыми (White flint) зерновками. Зерновки межсортовых гибридов можно было различить по их окраске. Опыление растений сорта White flint пыльцой растений сорта Yellow sweet приводит к образованию желтых плодов. В эксперименте выращивали смесь из индивидуумов обоих сортов. В начале эксперимента в среднем 35,8 % зерновок, образованных сортом White flint, были желтыми, а 46,7 % зерновок, образованных Yellow sweet, — белыми. Для скрещиваний в последующих поколениях высевали зерновки тех индивидуумов, которые имели самый низкий процент гибридных плодов. Таким образом проводили отбор против гибридизации. Через шесть поколений процент гибридных зерновок снизился соответственно до 4,9% (White flint) и 3,4 % (Yellow sweet). Уменьшение гибридизации стало возможным, потому что снизилось перекрывание сроков цветения обоих сортов. Признаки этого явления наблюдали и в природе. У Agrosüs capillaris известны генотипы, толерантные к тяжелым металлам. Популяции с этими генотипами часто непосредственно соседствуют с таковыми без устойчивости к тяжелым металлам, например, по краю рудничных отвалов. На медном руднике в Уэльсе удалось показать, что толерантные к тяжелым металлам растения на нормальной почве имеют пониженную приспособленность, а нетолерантные растения почти не способны расти на почвах с медью. Гибридное потомство между толерантными и нетолерантными индивидуумами имеет более низкую приспособленность, чем потомство от индивидуумов с одинаковыми генотипами. Это обусловливает (см. рис. 10.33) появление естественного отбора против гибридов. Наблюдали, что толерантные растения Agrostis capillaris начинают цвести примерно на неделю раньше нетолерантных индивидуумов. Эти различия в сроках цветения сохраняются и при культивировании в одинаковых условиях. Так как в целом сроки цветения значительно (на несколько недель) перекрываются, здесь можно предположить, по крайнем мере, начало эволюционного становления репродуктивной изоляции путем сдвига сроков цветения.
При очень сильном отборе против гибридов, т. е. при сильно сниженной приспособленности гибридных индивидуумов, возможны пара- или симпатрическое возникновение механизмов изоляции и тем самым пара- или симпатрическое видообразование. Правда, не ясно, существует ли в природе достаточно сильный отбор этого типа и быстрее ли он ведет к репродуктивной изоляции, чем генетический обмен к гомогенизации генетически дифференцированных популяций. По крайней мере, теоретическая возможность эволюции репродуктивной изоляции при пара- или симпатрии показывает, что репродуктивная изоляция не только бывает побочным продуктом эволюционной дивергенции, но и может возникать вследствие естественного отбора на изоляцию. Это важно и в тех случаях, когда начинают вторично контактировать неполностью репродуктивно изолированные близкородственные виды. В такой ситуации существующие несовершенные барьеры, препятствующие потоку генов, могут быть усилены естественным отбором (англ. reinforcement).
Обычно области гибридизации между видами известны как гибридные зоны. Такие гибридные зоны могут возникать либо вторично благодаря встрече ранее аллопатрически дифференцированных видов, либо, возможно, первично — в ходе пара- или симпатрического видообразования. И обоих случаях они функционируют как барьеры потоку генов, потому что пространственно изолируют друг от друга виды, например, отсеивают чужеродную пыльцу.
10.3.2.4. Генетика видовых различий
Аллопатрическое видообразование чаще всего рассматривают как постепенный процесс на основе наблюдений уменьшающегося фенотипического и генетического сходства и постепенно возрастающей репродуктивной изоляции между все менее близкородственными подвидами, видами и т. д. Эта гипотеза подразумевает, что и морфологические изменения — процесс постепенный. Генетические исследования видовых различий многократно показали (рис. 10.34), что изменчивость анализируемых признаков в любом выбранном поколении непрерывна; четкие классы признаков чаще всего выделить невозможно. Поэтому пришли к выводу, что на морфологические признаки влияют многие гены. Мутации некоторых из многочисленных генов должны были бы тогда приводить к мелким изменениям морфологического признака. Методы молекулярной генетики принесли новые результаты. Стало возможно идентифицировать и анализировать морфогенетические гены. Далее путем анализа распределения морфологических и молекулярных признаков, локализованных на генетической карте сцеплений, можно выявить, сколько генов и с каким относительным эффектом участвует в формировании фенотипа (см. 10.1.2.4).
Рис. 10.34. Теосинте (А — С, Z. mays subsp. parviglumis) и дикая форма культурной кукурузы (D — F, Z. mays subsp. mays). Различаются по архитектуре побеговой системы и расположению мужских и женских соцветий: А — теосинте: боковые побеги с верхушечными мужскими соцветиями; D— кукуруза; боковые побеги с верхушечными женскими соцветиями (различна также структура женских соцветий); В, С — теосинте; колоски в два ряда, 1 колосок в обертке; Е, F — кукуруза; многочисленные ряды колосков, 2 колоска в обертке. Эти различия определяются в основном двумя генами (teosinte branched 1, tb1 и teosinte glume architecture, tga1)
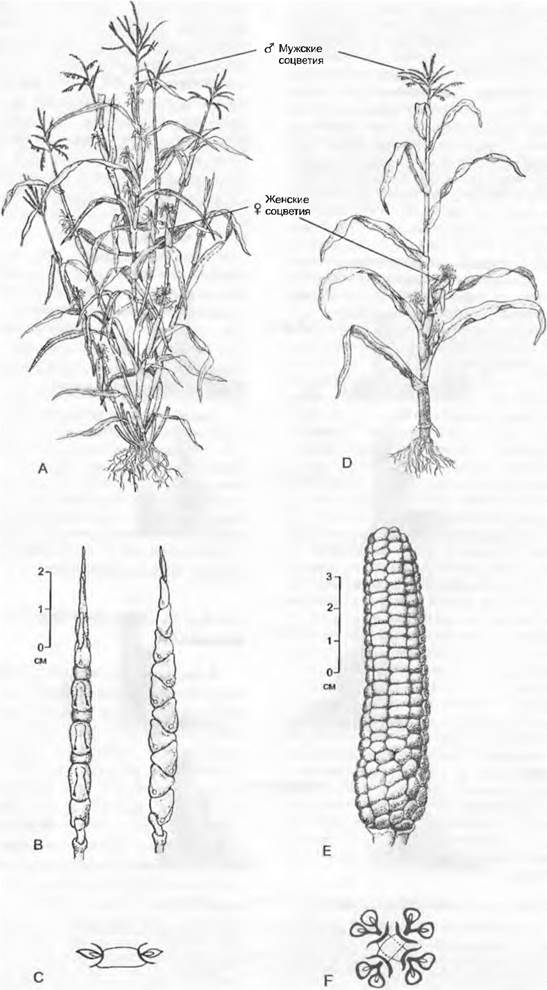
Особенно хорошо исследована в этом отношении кукуруза. Культивируемая кукуруза (Zea mays subsp. mays) происходит от теосинте (7 mays subsp. parviglumis) и, вероятно, впервые появилась около 10 тыс. лет назад. Важное различие между двумя этими подвидами — строение побеговой системы и расположение женских и мужских соцветий (см. рис. 10.34). У кукурузы главный побег заканчивается мужским соцветием, а боковые ветви первого порядка имеют очень короткие междоузлия и несут верхушечные початки. У теосинте главный побег также заканчивается мужским соцветием. Однако междоузлия боковых побегов первого порядка не укорочены, и побеги заканчиваются также мужскими соцветиями. Женские соцветия следует искать на верхушках боковых побегов второго порядка. Генетический и молекулярный анализ этих различий показал, что они обусловлены всего одним геном. Этот ген, известный у одного мутанта кукурузы как teosinte branched (tb 1), влияет на длину междоузлий боковых побегов и пол соцветий. Ген tb1 проявляется у теосинте только в пазушных почках боковых побегов первого порядка, из которых образуются женские соцветия. У мутантов культурной кукурузы, напротив, tb\ проявляется уже в пазушных почках главного побега. Это приводит к образованию укороченных боковых побегов с верхушечными женскими соцветиями. Экспрессия этого гена у кукурузы отчетливо сильнее, чем у теосинте. Эволюция здесь, похоже, проявляется главным образом в изменении генной регуляции, tb1 — регуляторный ген, влияющий, очевидно, на силу апикального доминирования.
Кукуруза и теосинте различаются и по структуре женских соцветий. У теосинте колоски расположены в два ряда. В каждом углублении на оси соцветия по одному колоску с очень жесткими колосковыми чешуями, охватывающими весь колосок. При созревании женские колоски разрываются. Напротив, у культурной кукурузы колоски расположены в многочисленных рядах. Углубления на оси початка мелкие, каждое содержит по два колоска. Колосковые чешуи маленькие, сравнительно мягкие и не охватывают колоски. При созревании колоски остаются неповрежденными. Удалось показать, что один ген (teosinte glume architecture (tga1)) влияет на жесткость колосковых чешуи, регулируя отложение в них кремния и лигнина, величину колосковых чешуй и глубину углублений на оси соцветия.
Другой пример большого эффекта генов дают виды губастика (Mimulus). Опыляемый шмелями Mimulus lewisii образует небольшое количество (около 0,5 мкл) очень концентрированного нектара. Опыляемый колибри М. сагdinalis, напротив, имеет много (около 40 мкл) сравнительно жидкого нектара. Это различие, по-видимому, регулируется преимущественно одним геном.
Такие различия показывают, что, несмотря на влияние на структуру многочисленных генов, морфологическая эволюция может идти путем глубоких преобразований благодаря изменениям регуляторного гена, занимающего более высокое положение в функциональной иерархии генов.
10.3.3. Гибридизация и гибридное видообразование
10.3.3.1. Гибридизация
При всей значимости репродуктивной изоляции для видообразования многие виды растений репродуктивно не изолированы друг от друга и могут гибридизировать. Понятие «гибридизация» здесь следует применять к скрещиванию между видами, но в расширенном смысле его можно использовать для обозначения скрещивания между генетически дифференцированными популяциями, подвидами и другими группировками внутри вида.
Частоту скрещивания между видами в природе можно наблюдать на различных уровнях. С одной стороны, анализ пяти хорошо известных флор умеренного климата показал, что гибридны от 5,8 % («межгорная» флора Кордильер, США) до 22% (Британские острова) от общего числа видов высших растений этих флор. С другой стороны, анализ смешанных популяций гибридизирующих видов показал, что гибриды составляют от <1 % (например, Senecio vernalis у S. vulgaris) до 31% (например, у Quercus) индивидуумов этих популяций.
Гибридизация часто бывает асимметричной, когда она успешна только в тех случаях, если два вида выступают в качестве материнского и отцовского растений в определенной комбинации. Например, скрещивание между двумя видами примул — Primula vulgaris и Р. veris — бывает успешным, если только в качестве материнского растения выступает Р. veris.Очень часто скрещивание между самосовместимым и самонесовместимым видами приводит к гибридному потомству лишь в тех случаях, когда материнским растением был самосовместимый вид.
Поскольку частота гибридизации между видами зависит от мощности пре- и постзиготических изоляционных барьеров, она связана и с относительной частотой гибридизирующих видов в их смешанной популяции. Частота гибридизации обычно увеличивается, если один из двух видов встречается в значительно меньшем числе. Это можно объяснить среди прочих причин тем, что вероятность опыления чужеродной пыльцой вида, представленного лишь немногими индивидуумами, в такой ситуации выше, чем при равной численности обоих видов.
Наконец, наличие гибридизации в значительной мере коррелирует с нарушениями окружающей среды либо вследствие естественных причин (например, климатические изменения в четвертичном периоде), либо под воздействием человека. Эффект нарушения состоит в том, что он допускает взаимные встречи видов, географически или экологически разделенных в ненарушенной среде чем повышается вероятность появления гибридных индивидуумов; если в биотопе, заселенном родительскими видами, гибриды чаще всего оказываются угнетенными, то в так называемом гибридном местообитании или во вновь заселяемых биотопах они могут иметь преимущество перед родительскими видами.
Гибриды по морфологическим признакам промежуточны между родительскими видами, но иногда имеют и новые признаки, т. е. (рис. 10.35) не встречающиеся у родительских видов. Новые признаки у гибридов могут получиться как благодаря новым комбинациям генов, не встречающимся у родительских видов, так и благодаря гетерозиготности отдельных генных локусов.
Распределение морфологических признаков предположительно гибридной популяции (или гибридного вида) в сравнении с таковым родительских видов представлено на диаграмме (см. рис. 10.35). Оно может быть важным инструментом для подтверждения предполагаемого гибридного происхождения. Гибридные индивидуумы часто содержат вторичные вещества, характерные для обоих родительских форм (рис. 10.36), но нередко и в этих случаях встречаются новые вещества. На уровне генетических признаков свойства гибрида зависят и от хода наследования рассматриваемого признака. В то время как у гибридов, по крайней мере первого поколения, при бипарентальном наследовании ДНК находят признаки ДНК обоих родительских видов, гибрид при наследовании ДНК по материнской или отцовской линии может обладать ДНК только одного из родителей.
Рис. 10.35. Диаграмма гибридологического анализа. Экспериментальное скрещивание диплоидных видов Achillea setacea (Ps) и A. asplennfolia (Ра) (по F. Ehrendorfer).
Разные индивидуумы F1от реципрокных скрещиваний (s х а, а х s) обведены пунктирной линией, родительские индивидуумы — штриховой линией. Все остальные точки представляют собой субвитальное F2. Спонтанно возникший аллотетраплоидный индивидуум показан серым цветом
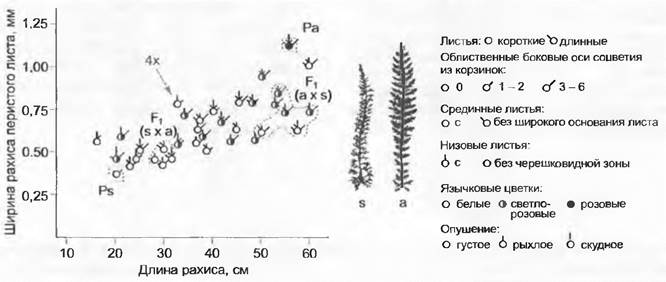
Рис. 10.36. Возникновение и анализ полиплоидного комплекса у папоротников (североамериканские виды Asplénium). При подсчетах хромосомных чисел и пар конъюгирующих хромосом у гибридов установлены формулы геномов: диплоидные исходные виды A. platyneuron (PP), A. rhizophyllum (RR) A. montanum (ММ), ди-, три- или тетраплоидные гибриды: RP, RMM, RMP и RMPM, а также аллотетраплоидные дочерние виды A. ebenoicles (RRPP), A. pinnatifidum (RRMM) и A. bradleyi (PPMM); история их возникновения подтверждена морфологией, сравнительной фитохимией (фенольные соединения: ксантоны, по результатам двумерной бумажной хроматографии), а также анализом образцов изо- и аллозимов

В ходе дальнейшей эволюции гибридов может наблюдаться гомогенизация последовательности ДНК в направлении одного из родительских видов, как, например, обычно используемого в систематике внутреннего транскрибируемого участка рибосомальной ДНК (ITS). В результате через несколько поколений у гибридов возможно обнаружить ITS-последовательность только одного из родителей. Этот феномен — проявление так называемой согласованной эволюции, которая особенно характерна для повторяющихся последовательностей и включает также гомогенизацию многочисленных повторов.
Часто гибриды, по крайней мере между близкородственными видами, отличаются от родителей более мощным ростом. Это явление, хорошо известное как гетерозис, отражает гетерозиготность многочисленных локусов у гибридных индивидуумов.
Данную особенность используют в сельском хозяйстве. Урожайность многих культурных растений (например, кукурузы, сахарной свеклы) можно повысить, используя посевной материал, полученный от скрещивания «чистых» (инбредных) линий и дающий сильный эффект гетерозиса. Получение такого посевного материала облегчает использование одной из родительских линий со стерильной пыльцой.
Фертильность гибридов совершенно не связана с гетерозисом. В зависимости от мощности постзиготических изолирующих барьеров гибридные индивидуумы F1 нередко демонстрируют пониженную фертильность. Но так как фертильность разных гибридных индивидуумов, получившихся в результате одного и того же скрещивания, вариабельна, существует возможность того, что отдельные гибридные индивидуумы мало и совсем не отличаются по фертильности от родительских индивидуумов. В то время как приспособленность гибридов в местообитании родительских видов чаще всего ниже, чем у родительских растений, приспособленность гибридов в «гибридном» местообитании может оказаться даже выше, чем у их родителей.
Обычно существование гибридов ограничено во времени и пространстве, но существует возможность того, что они становятся исходным пунктом дальнейшей эволюции и возникновения новых видов. Предпосылкой этого является неполная стерильность гибридных индивидуумов. Для успешного появления гибридного потомства важно быстрое восстановление неограниченной способности к размножению. Оно может происходить или без изменения числа хромосом (гомоплоидное гибридное видообразование и интрогрессивная гибридизация), или в ходе полиплоидизации (аллополиплоидное гибридное видообразование). Наконец, полностью сте
рильные гомоплоидные или полиплоидные гибриды могут размножаться апомиктически.
10.3.3.2. Гомоплоидное гибридное видообразование
Условием восстановления полной фертильности гибридов без изменения числа хромосом (гомоплоидное гибридное видообразование) является рекомбинация в гибридном потомстве. Для этого необходимо, чтобы гибридные индивидуумы могли размножаться либо путем самооплодотворения, либо благодаря скрещиванию друг с другом или возвратному скрещиванию с родительскими видами. При отборе на повышение фертильности результат рекомбинации должен состоять в том, чтобы гибридное потомство становилось гомозиготным по таким факторам, по которым различаются родительские виды и которые как элементы постзиготических механизмов изоляции вызывают пониженную фертильность гибридов. Этими факторами являются генные или чаще хромосомные мутации, по которым различаются родительские виды (см. 10.1.2.2). Поэтому гомоплоидное видообразование часто называют также рекомбинационным видообразованием. Соответственно частоте хромосомных мутаций у гибридизирующих видов гомоплоидное видообразование обычно связано с быстрым изменением структуры хромосом благодаря последующим хромосомным мутациям. Эта хромосомная эволюция у гибридного потомства одновременно приводит к установлению репродуктивной изоляции гибридов от родительских видов. Далее для существования фертильного гибридного потомства важно, чтобы для него нашлось подходящее местообитание.
Процесс гомоплоидного видообразования в связи с хромосомными мутациями и заселением нового местообитания особенно хорошо описан у подсолнечников (Helianthus) в Северной Америке. От гибридизации Helianthus annum, чаще всего обитающего на тяжелых глинистых почвах, и Н. petiolaris с песчаных почв возникли три гибридных вида: Н. anomalus, Н. deserticola и Н. paradoxus. В противоположность родительским видам Н. anomalus и Н. deserticola заселяют крайне сухие почвы, а Н. paradoxus— влажные солонцы. Сопоставление последовательности генов в хромосомах Helianthus annuus и Н. petiolarls как родительских видов и Н. anomalus как гибридного потомка показало, что родительские виды с гаплоидным числом хромосом 17 различаются по меньшей мере десятью хромосомными мутациями (три инверсии, семь транслокаций). Гибридный потомок Н. anomalus имеет в шести хромосомах такое же расположение генов, как у обоих родительских видов, в четырех других хромосомах гены расположены либо как у одного, либо как у другого родительского вида, а семь из 17 хромосом отличаются от хромосом обоих родителей (рис. 10.37). Эти преобразования, по-видимому, могли произойти очень быстро. Экспериментальное получение гибридов между Н. annuus и Н. petiolaris с последующим отбором на фертильность привело к ее росту от <10 % у гибридов F1 к почти полной фертильности после пяти поколений. Структура хромосом этих искусственных гибридов была удивительно похожа на хромосомы встречающегося в природе Н. anomalus.
Рис. 10.37. Рекомбинация у гомоплоидного гибридного вида. При гаплоидном числе х = 17 подсолнечники Helianthus annuus и Н. petiolaris различаются по меньшей мере 10 хромосомными мутациями (3 инверсии, 7 транслокаций). Гомоплоидный гибридный потомок Н. anomalus в 6 хромосомах (А — F) имеет такое же расположение генов, как у обоих родительских видов, 4 хромосомы (L/M, N, Т U) демонстрируют расположение генов как у одного из родителей, а остальные 7 хромосом отличаются от обоих родительских видов

Быстрое восстановление высокой фертильности показано и в селекционных экспериментах с гибридами, например, Elymus и Cilia. Наряду с Helianthus гомоплоидное видообраование хорошо документировано также в родах iris, Paeonia. Pinus и Stephanomeria. Удалось показать, что в возникновении Iris nelsonii принимали участие даже три вида (I. fulva, I. hexagona, I. brevicaulis).
10.3.3.3. Интрогрессивная гибридизация
В зависимости от относительной частоты самооплодотворения (насколько она вообще возможна) у гибридов, их скрещивания друг с другом и возвратного скрещивания с родительскими видами гибридное потомство в дополнение к случайно появившимся новым признакам будет иметь более или менее промежуточное строение, т. е. проявит большее или меньшее сходство с родительскими видами. Регулярное или по крайней мере сравнительно частое возвратное скрещивание гибридного потомства с одним из родительских видов может привести к тому, что только относительно немногие признаки одного вида постоянно переходят к другому виду. Этот процесс, названный интрогрессивной гибридизацией, или интрогрессией, в принципе соответствует традиционной селекции растений, когда путем скрещиваний пытаются передать культурным растениям желаемые особенности дикорастущих видов (например, устойчивость к грибным заболеваниям).
Интрогрессия выявлена, например, у крестовника обыкновенного (Senecio vulgaris) в Великобритании (рис. 10.38). В 1875 г. у этого вида, в норме не имеющего язычковых цветков в корзинках (var. vulgaris), была описана разновидность (var. hibernicus) с короткими язычковыми цветками. Путем экспериментального ресинтеза и использования молекулярных признаков удалось показать, что данный признак возник благодаря интрогрессивной гибридизации между занесенным в Великобританию в XVIII столетии S. squalidus с крупными язычковыми цветками и S. vulgaris. S. squalidus и сам является гомоплоидным гибридным видом, возникшим из сицилианских S. aethnensis и S. chrysanthemifolius.
Рис. 10.38. Интрогрессивная гибридизация. Короткие язычковые цветки в корзинках Senecio vulgaris var. hibernicus (С) получились благодаря интрогрессивной гибридизации между S. vulgaris var. vulgaris (В) без язычковых цветков и S. squalidus (А) с длинными язычковыми цветками
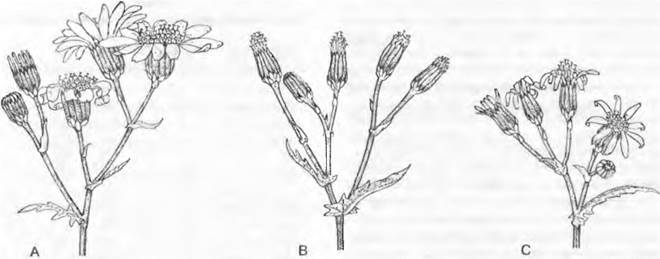
Как показывает пример широко распространенной в Великобритании var. hibemicus, интрогрессия не ограничена узкими пространственными пределами.
Сравнение пластидных и ядерных ДНК привело к выводу, что интрогрессия пластидных ДНК встречается значительно чаще, нежели интрогрессия ядерных ДНК. Так, нередко обнаруживают, что индивидуумы одного вида обладают пластидным геномом другого вида, а в их ядерной ДНК нет признаков гибридизации. Этот феномен также известен как хлоропластный захват («chloroplast capture).
Из 141 исследованного в Южной Калифорнии индивидуума Helianthus petiolaris 137 обладали пластидным геномом Н. annuus. Только два индивидуума Н. petiolaris имели признаки Н. annuus и в ядерном геноме. Так как Н. petiolaris был занесен в Калифорнию всего около 50 лет назад, этот пример иллюстрирует и потенциально высокую скорость интрогрессивной гибридизации.
Успешное появление гомоплоидных гибридных видов или возникающих путем интрогрессии новых форм одного вида ос
новано преимущественно на том, что гибриды имеют повышенную генетическую изменчивость благодаря комбинации родительских геномов. Рекомбинация этих вариаций в последующих гибридных поколениях при гомоплоидном гибридном видообразовании может привести к возникновению новых признаков.
Однако гибридизация и интрогрессия ведут также и к снижению репродуктивной изоляции. Это позволяет S. vulgaris var. hibernicus, возникшей путем интрогрессии между Seneciovulgaris и S. squalidus, легче скрещиваться с S. squalidus, чем с незатронутой гибридизацией var. vulgaris, что возможно объяснить большим генетическим сходством var. hibernicus с S. squalidus, генетический материал которого она инкорпорировала, чем сходством var. vulgaris с этим видом. Уменьшение репродуктивной изоляции может привести в конце концов к слиянию двух видов путем гибридизации. Показано, что редкому Канарскому локальному эндемику Argyranthemum coronopifolium как виду угрожает исчезновение вследствие гибридизации с быстро распространяющимся на Канарских островах под влиянием человека A. frutescens. Таким образом возникает важный аспект охраны редких видов — снижение гибридизации между редкими и обычными видами.
10.3.3.4. Аллополиплоидия
Фертильность гибридов, чаще всего пониженная благодаря постзиготическим механизмам изоляции, может восстановиться путем полиплоидизации. Для пояснения обозначим разные гаплоидные геномы гибридизирующих видов как А и В (или А и A’ при менее выраженных различиях): диплоидные гибридные индивидуумы имеют тогда геномный состав АВ (или АА’). Из-за недостаточной гомологии хромосом вследствие генных либо хромосомных различий образование бивалентов в мейозе более или менее нарушено, что и снижает фертильность (см. 2.2.3.7). Если в ходе гибридизации благодаря слиянию нередуцированных гамет непосредственно или через анортоплоидную промежуточную стадию (см. 10.1.2.3) происходит удвоение генома, то получаются тетраплоидные гибриды с геномным составом ААВВ или ААА’А’, у которых в мейозе каждая хромосома находит себе гомологичного партнера для конъюгации. Это приводит к правильному образованию бивалентов и чаще всего имеет следствием полное восстановление фертильности. Данная модель особенно убедительна, если гибридизирующие виды различаются хромосомными мутациями.
Воздействие полиплоидизации на фертильность более или менее стерильных диплоидных гибридов доказано многократно. В качестве примера можно привести еще раз гибрид между Prímula floribunda и Р. verticillata (P. х kewensis), у которого соматическая полипловдизация обычного стерильного индивидуума привела к образованию единственного фертильного соцветия. Регулярное образование бивалентов у полиплоидных гибридных растений является, однако, функцией не только гомологии между хромосомами от одного родителя, но и отсутствия гомологии между хромосомами разных родителей. Путем последовательной элиминации 21 пары различающихся хромосом аллогексаплоидной (2т = 6х = 42) мягкой пшеницы (Triticum salivum) сталось показать, что при отсутствии 5-й хромосомы В-генома регулярное образование бивалентов нарушается и имеет место мультивалентность. Это позволяет сделать вывод, что образование бивалентов также находится под генетическим контролем, причем в данном случае важные для этого гены локализованы в названной хромосоме.
Полиплоидные гибриды в известной мере репродуктивно изолированы от родительских видов вследствие измененного числа хромосом. Возвратное скрещивание, например, тетраплоидного (4х) гибрида с одним из диплоидных родителей (2х) ведет к образованию триплоидных индивидуумов (Зх), мейоз у которых существенно нарушен из-за невозможности конъюгации одного из трех гаплоидных геномов и которые в соответствии с этим имеют пониженную фертильность.
Однако поток генов между индивидуумами с разной плоидностью не исключен полностью, так как, например, триплоидные растения все же образуют небольшое число диплоидных гамет (см. 10.1.2.3), которые при слиянии с диплоидными гаметами тетраплоидных гибридов могут привести к фертильному потомству. В качестве одного из многих примеров потока генов между растениями различной плоидности можно сослаться на интрогрессивную гибридизацию между Senecio vulgaris и S. squalidus, поскольку S. squalidus — диплоид (2n = 20), a S. vulgaris — тетра- плоид (2n = 40).
Считается, что шансы возникших полиплоидных гибридов могут сохраниться, как и у гомоплоидных гибридов они увеличиваются, если в их распоряжении оказываются нарушенное «гибридное» местообитание либо новый биотоп для заселения. Часто это можно заметить в географическом распространении диплоидных и полиплоидных видов одного рода.
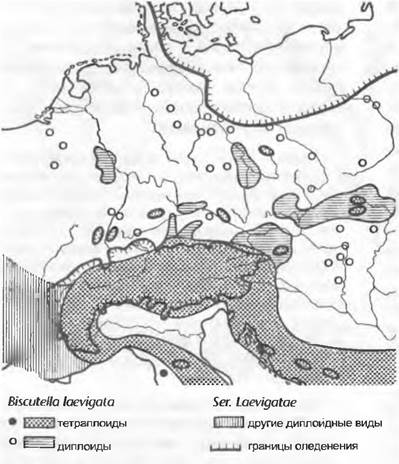
Диплоидные виды и подвиды Biscutella ser. Laevigaíae произрастают далеко за пределами области, подвергшейся оледенению в четвертичном периоде, в то время как тетраплоидные таксоны можно встретить главным образом внутри этой области (рис. 10.39). Эффект естественного нарушения местообитаний из-за постоянно меняющегося в четвертичном периоде положения ледника состоял, с одной стороны, в том, что климатические изменения допускали тогда контакты гибридизирующих таксонов, а с другой стороны, в том, что отступление ледника освобождало огромные пространства, которые могли заселять новообразующиеся полиплоидные таксоны. Аналогичный пример дают североамериканские ирисы Iris virginica (2n = 72), I. setosa subsp. interior (2n = 72) и их аллополиплоидный гибрид I. versicolor (2n = 108). В то время как /. virginica и /. setosaвстречаются в далеких областях, не подвергавшихся оледенению (юго-восток Северной Америки и Аляска), /. versicolor растет почти исключительно в прежде покрытых ледником областях северо-востока Северной Америки.
Рис. 10.39. Распространение диплоидных и тетраплоидных форм Biscutella ser. Laevigatae в Центральной Европе. Диплоиды предпочли неоледеневавшие области в эпоху вюрмского оледенения, тетраплоиды можно найти также в подвергшихся сильному оледенению Альпах
Уже давно известный факт, что частота полиплоидных растений возрастает с увеличением географической широты (доля полиплоидов в скандинавской флоре составляет 56 — 72 %, а в североафриканской, средиземноморской и Канарской флорах — 23 — 34%), лучше всего можно объяснить изменениями климата в палеогеннеогеновый и преимущественно в четвертичные периоды. Климатически обусловленные сдвиги ареалов и возникновение незаселенных территорий привели, как и в случае с американскими Biscutella, к появлению полиплоидных гибридных видов.
На возникновение полиплоидного гибридного вида может указывать комбинация родительских признаков, выявляемая морфологическим, фитохимическим, кариологическим и молекулярным методами.
Особенно показательны для кариологических признаков методы видоспецифичной дифференциальной окраски хромосом. К примеру, хромосомные препараты у полиплоидного гибридного вида можно обработать видоспецифическими зондами ДНК, связанными с флуоресцентными красителями, и сделать таким образом узнаваемыми хромосомы его родителей (рис. 10.40). Эта техника известна и как метод хромосомных отпечатков («chromosome painting») или FISH (Fluorescence In Situ Hybridisation). Гибридизацию геномных ДНК на клеточных препаратах в целом называют GISH-техникой (Genomic In Situ Hybridisation).
Рис. 10.40. Геномная гибридизация in-situ («genome painting») соматических хромосом у аллотетра- плоидного табака (Nicotiana tabacum, 2n = 48): А — фото; В — поясняющий рисунок. Специфическое связывание ДНК и флуорохромная окраска позволяют различить хромосомные наборы обоих диплоидных родительских видов, белым показан N. otophora(2n = 24), серым — N. sylvestris (2n = 24). Пары хромосом 1 — 3 имеют смешанную окраску: здесь произошла транслокация между хромосомами родительских видов
Кроме этих приемов, можно скрещивать полиплоидные гибридные виды и с их предполагаемыми диплоидными родительскими видами. В мейозе у такого триплоидного индивидуума в самом благоприятном случае удается наблюдать, что два из трех имеющихся гаплоидных хромосомных наборов образуют биваленты, а третий хромосомный набор в конъюгации участия не принимает. Это может означать, что использованный диплоидный вид действительно был родителем гибридного вида. Если образования бивалентов не происходит, то использованный в скрещивании диплоидный вид можно исключить из числа предполагаемых родителей. У триплоидного гибрида между аллотетраплоидным папоротником Asplénium pinnatifîdum с генной формулой RRMM и одним из диплоидных родительских видов (A. montanum: ММ) можно наблюдать конъюгацию хромосом только М-генома. Однако возможности этой техники анализа полиплоидных гибридов ограничены, так как конъюгация хромосом не только зависит от их гомологии, но и находится под генетическим контролем, а для отсутствия бивалентов чаще всего требуется сильная дивергенция хромосом обоих родительских видов.
Наконец, важным средством доказательства является экспериментальный ресинтез полиплоидного гибридного вида из предполагаемых родительских видов. Первый такой ресинтез был осуществлен в 1930 г. А. Мюнтцингом. Мюнтцинг предположил, что пикульник обыкновенный (Galeopsis tetrahit) мог возникнуть как полиплоидный гибридный вид (2n = 4х = 32) из пикуль- ника пушистого (G. pubescens; 2n = 2х =16) и пикульника красивого (G. speciosa; 2n = 2х = 16). Для доказательства этой гипотезы он скрестил оба последних вида. Самоопыление почти полностью стерильных гибридов дало потомство, в котором был обнаружен единственный триплоидный индивидуум. Скрещивание этого растения с G. pubescens привело к появлению тетраплоидных растений, морфологически очень похожих на G. tetrahit и успешно скрещивавшихся с данным видом.
В других случаях был выбран прямой путь экспериментального ресинтеза. Так табак деревенский, или махорка (Nicotiana rustica; 2n = 4х = 28), был получен в результате полиплоидизации с помощью колхицина гибрида между N. раniculata и N. undulata (оба имели 2n = 2х = 24), а рапс (Brassica napus-, 2n = 2х = 38) — в результате полиплоидизации гибрида между капустой (В. olerácea; 2n = 2 х = 18) и репой (В. rapa; 2n = 2х = 20).
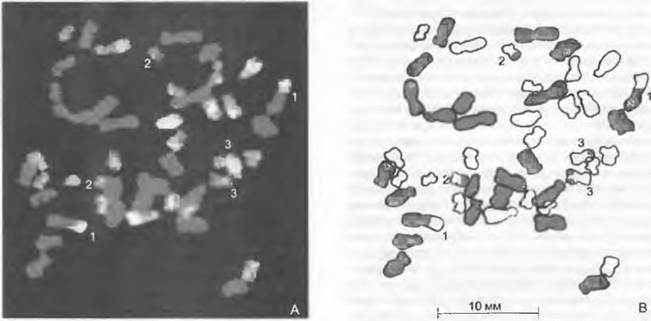
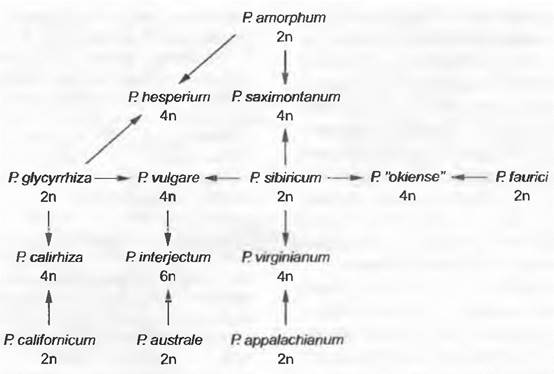
Из многочисленных хорошо исследованных примеров возникновения полиплоидных гибридных видов следует еще кратко описать папоротник многоножку (Polypodium), пшеницу мягкую (Triticum aestivum) и козлобородник (Tragopogón). Использовав очень много признаков, удалось показать, что семь диплоидных видов Polypodium принимали участие в возникновении шести тетраплоидных и одного гексаплоидного вида (см. рис. 10.40, рис. 10.41). При этом тетра плоидная многоножка обыкновенная (Р. vulgare) возникла вследствие гибридизации диплоидных P. glycyrrhiza и Р sibiñcum и в свою очередь вместе с диплоидной P. australe участвовала в возникновении гексаплоидной P. interjectum. Благодаря анализу у наследуемой по материнской линии пластидной ДНК удалось надежно показать, что некоторые виды Polypodium (Р. са- lirhiza, P. hesperium, P. virginianum) содержат пластидную ДНК обоих родителей и потому, очевидно, возникали по меньшей мере дважды.
Рис. 10.41. Полиплоидный комплекс в роде Polypodium 7 диплоидных видов рода Polypodium участвовали в образовании 6 тетраплоидных и 1 гексаплоид- ного вида. Тетраплоидная многоножка обыкновенная (Р. vulgare) возникла в результате гибридизации диплоидных Р- glycyrrhiza и Р sibiricum. Сравнительный анализ признаков, унаследованных от материнской формы и от обоих родителей, показал, что некоторые виды возникали по крайней мере дважды (Р. calirhiza, Р. hesperium, Р. virginianum)
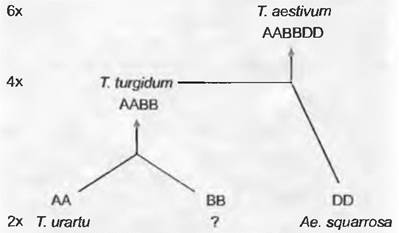
Гексаплоидная (2n = 6х = 42) мягкая пшеница возникла из трех разных видов, из которых, правда, сегодня можно назвать только два (рис. 10.42). Вначале Тriticum urartu (геномная формула АА) гибридизировала с неизвестным видом (ВВ) Возникшая в результате Тriticum turgidum (ААВВ) в конце третьего тысячелетия до нашей эры вновь гибридизировала в качестве материнского организма с Aegilops squcirrosa (DD), дав современную мягкую пшеницу с геномным составом AABBDD. Археологические находки четко показывают, что люди культивировали уже и диплоидные и тетраплоидные виды — соответственно, однозернянку и эммер (рис. 10.42).
Рис. 10.42. Возникновение гексаплоидной пшеницы мягкой (2n = 6х = 42) путем аллополиплоидии. Гибридизация Тriticum urartu (формула генома АА) с неизвестным видом (ВВ) привела к Тгiticum turgidum (ААВВ), которая в качестве материнского вида после скрещивания с Аеgilops squarrosa (DD) и дала современную мягкую пшеницу с геномным составом ААВBDD
Как и мягкая пшеница, многие другие культурные растения также имеют полиплоидное и гибридное происхождение. К ним относятся, например, уже названные рапс (Brassica), табак (Nicotiana), овес (Avena), виноград (Vitis) и т. д. Полиплоидия у культурных растений, впрочем, не более обычна, чем у родственных им дикорастущих видов. Это можно объяснить, вероятно, тем, что нарушение естественной растительности и культивирование растений человеком аналогично эффекту климатических изменений в четвертичном периоде привели к тому, что географически и экологически изолированные в природе виды встретились и могли гибридизировать, причем уже на ранней земледельческой стадии возделывание более или менее освобожденных от конкуренции культурных растений допускало выживание полиплоидных гибридных видов.
В роде козлобородник (Tragopogón) три европейских вида — Т. dubius, Т. porrifolius и Т. pratensis, которые были занесены в Северную Америку, от скрещивания между соответственно Т. dubius и Т. porrifolius и Т. dubius и Т. pratensis возникли тетраплоидные гибридные виды Т. mirus и Т mis- cellus. Путем анализа как ядерной, так и пластидной ДНК было показано, что оба тетраплоидных вида подобно видам Polypodium возникали повторно. Так, Т. miscellus в одном случае имел в качестве материнского растения Т. dubius, а во всех других — Т. pratensis. Повторное возникновение ал лополиплоидных видов удавалось доказать неоднократно.
Тот факт, что аллотетраплоидные Т. mirus и Т. miscellus возникли только в Америке, но не на своей европейской родине, еще раз показывает, что для выживания гибридных видов требуется измененная окружающая среда. Вероятно, отсутствие гибридных видов в Европе связано не с тем, что они там не возникали, а с тем, что они не смогли там закрепиться.
Как и диплоидные гибридные растения, полиплоидные гибриды проявляют большую генетическую изменчивость. Она выражается в повышенной гетерозиготности (в зависимости от аллельной конституции родительских видов) вследствие большого числа аллелей в локусе и большого числа гетерозиготных локусов, а также в образовании новых комбинаций генов.
В то время как у диплоидных гибридов гетерозиготность преходящая вследствие сегрегации аллелей в чреде поколений, у полиплоидов гетерозиготность может фиксироваться. Это происходит, когда гомологичные локусы содержат разные аллели в хромосомах, полученных от обоих различающихся родителей, когда соответствующие родительские хромосомы не образуют биваленты и, соответственно, отсутствует сегрегация геномов. Удвоение всех общих обоим родителям генов при полиплоидизации позволяет, так же, как и удвоение генов при хромосомных мутациях (см. 10.1.2.2), изменить функцию одного из удвоенных генов в последующей эволюции полиплоидов. С другой стороны, известно, что многократно повторяющиеся гены могут утратить свою функцию. Этот феномен известен как «молчащие гены» (gene silencing). Хота у полиплоидных гибридных растений чаще всего, по-видимому, конъюгируют гомологичные хромосомы одного родительского генома, не исключена и конъюгация гомологичных хромосом, унаследованных от разных родителей, и таким путем — рекомбинация между разными родительскими геномами (межгеномная рекомбинация). Показано, что геномы поколений F2 и F5, полученных от самооплодотворения полиплоидных гибридов F1, искусственно созданных в ходе скрещиваний в разных комбинациях Brassica rapa, В. nigra и В. olerácea, различаются по 38 — 96 признакам. Сделан вывод, что эти различия, установленные молекулярными методами, обусловлены перестройками хромосом. Частота геномных изменений в этом примере увеличивалась с возрастанием различий между родительскими геномами. Методом хромосомных отпечатков удалось показать межгеномные транслокации и у табака (Nicotiana) (см. рис. 10.40). Как и у гомоплоидных гибридов, у полиплоидных гибридов рекомбинация высвобождает дополнительную генетическую изменчивость.
Этим увеличением генетической изменчивости, вероятно, и обусловлен очевидный успех полиплоидных гибридных видов в эволюции почти всех групп растений.
На примере Polypodium и Tragopogón было показано, что один диплоидный вид может участвовать в образовании нескольких полиплоидных гибридных видов. Благодаря этому разные полиплоидные гибридные виды имеют общий геном, полученный от одного и того же диплоидного вида, поэтому репродуктивные барьеры между аллополиплоидами обычно слабее, чем между исходными диплоидными видами. Вследствие гибридизации аллополиплоидов друг с другом или с диплоцдами образуются полиплоидные ряды.
Примеры таких рядов — упомянутые многоножка (Polypodium: 2х, 4х. 6х) и пшеница (Triticum: 2х, 4х, 6х). Еще более длинные полиплоидные ряды обнаружены, например, у подмаренника (Galium anisophyllum: от 2х до 10х) или в роде щавель (Rumex: от 2х до 20х).
Возраст полиплоидов оценивают в зависимости от относительной частоты диплоидных и полиплоидных видов в полиплоидных рядах. Если в обширном круге родственных форм лишь немногие виды полиплоидные, то говорят о неополиплоидии. Если же, напротив, имеется много полиплоидов порой с очень большим числом хромосом при совершенном отсутствии диплоидных форм, то такой случай называют палеополиплоидией.
Крайний случай палеополиплоидии — род ужовник (Орhioglossum), в котором у О. reticulatum известен хромосомный набор 2n =1440 (96-плоид). Магнолиевые (Маgnoliасеае) с гаплоидным набором хромосом х = 19 также считают палеополиплоидами. Нео- и палеополиплоидии связаны в непрерывный ряд промежуточными состояниями плоидносги и значениями хромосомных чисел.
Как гомоплоидное, так и полиплоидное гибридное видообразование начинается с гибридизации видов, совместно встречающихся в одной области. И если впоследствии гибридные виды благополучно закрепляются, заселяют местообитание, отличающееся от родительских местообитаний, и наконец иную область, то само гибридное видообразование происходит симпатрически. Этим гибридное видообразование отличается от наиболее обычного аллопатрического видообразования путем эволюционной дивергенции в географически разделенных популяциях. Симпатрия родительских видов и гибридного потомства возможна благодаря тому, что между ними возникает сильная (хотя и не всегда полная) репродуктивная изоляция вследствие рекомбинации в случае гомоплоидного гибридного видообразования и вследствие изменения числа хромосом в случае полиплоидного гибридного видообразования. Полиплоидное гибридное видообразование особенно примечательно тем, что оно более или менее прерывисто и потенциально может произойти за одно поколение. Как и гомоплоидному видообразованию, ему приписывают высокую скорость. Это еще одно отличие от постепенной эволюцион ной дивергенции географически разделенных популяций.
Очевидно, что гибридизация с ее различными возможностями стабилизации гибридного потомства — весьма обычный процесс эволюционного изменения и видообразования в растительном мире. Возможность широкого участия гибридизационных процессов в эволюции растений (такая эволюция называется также сетчатой) всегда следует принимать во внимание при реконструкции филогенетических отношений.
Гибриды с пониженной фертильностью могут сохраниться благодаря агамоспермии или вегетативному размножению. Полиплоидные и обычно анортоплоидные (например, Зх, 5х) хромосомные числа имеют преимущественно формы с гаметофитной агамоспермией (см. 10.1.3.3). Уже не раз предполагали, что агамоспермные виды возникли как более или менее стерильные гибриды.
В роде рябина (Sorbus) есть три широко распространенных амфимиктических вида — ария (S. aria), рябина обыкновенная (S. aucuparia) и глоговина (S. tominalis) с диплоидным хромосомным числом 2n = 2х =34. В результате гибридизации этих трех видов в различных комбинациях и частично при участии нередуцированных гамет возникли многочисленные агамоспермные группы видов. Так, эндемичная для западной Англии S. bristolensis с триплоидным хромосомным числом 2n = Зх = 51 происходит, по-видимому, от скрещивания S. torminalis х S. aria. По-видимому, из этого же скрещивания следует выводить и агамоспермный, но диплоидный сборный вид S. latifolia, состоящий из множества узко распространенных мелких видов.
Признают также гибридогенное возникновение форм мятлика альпийского (Роа alpina), представленных амфимиктическими диплоидами и тетраплоидами, и апомиктическими (живородящими или агамоспермными) полиплоидами и анеуплоидами (2n = 31 — 61), агамосперм- ной лапчатки Нойманна (сборный вид Potentilla neumanniana; 4х — 12х) и многих агамоспермных видов из родов малина (Rubus), манжетка (Alchemillа), ястребинка (Hieradum) и одуванчик (Taraxacum). Так как агамоспермия сравнительно редко полностью облигатна (см. 10.1.3.3), половой процесс, происходящий время от времени у преимущественно агамоспермных гибридогенных видов, может привести благодаря дальнейшей гибридизации к чрезвычайно сложным и динамично изменяющимся комплексам форм. Особенно яркий пример этого — малина и ежевика (Rubus).
Стерильные гибриды могут сохраняться и благодаря вегетативному размножению. Пример — гибрид (Circaea х intermedia) между двулепестниками альпийским (С. alpina) и парижским (С. lutetiana), который несмотря на почти полную стерильность широко распространился благодаря вегетативному размножению.
Гибрид между европейской Spartina maritima (2n = 60) и завезенной из Северной Америки S. alternifolia (2n = 62) закрепился, с одной стороны, благодаря вегетативному размножению без изменения числа хромосом (S. х townsendit), а с другой стороны, благодаря полиплоидии как фертильный вид (S. anglica; 2n =120, 122). У чистяка (Ranunculus ficaria) диплоидные формы размножаются половым путем, но триплоидные и тетраплоидные формы — посредством образующихся в пазухах листьев выводковых клубеньков.