Генетика з основами селекції - М.П.Мигун - 2008
РОЗДІЛ II. Матеріальні основи та молекулярні механізми спадковості
2.4. Нуклеїнові кислоти - носії і гаранти реалізації генетичної інформації
Генетична інформація. про всі властивості та ознаки організму - будову, фізіологічні особливості та процеси розвитку - записана на Молекулах генетичних нуклеїнових кислот (гНК).
Дискретна одиниця спадковості - ген - є не чим іншим, як фрагментом молекули гНК.
Не всі існуючі в клітині нуклеїнові кислоти - генетичні. Так, наприклад, інформаційна РНК (іРНК), транспортні РНК (тРНК), рибосомнІ РНК (рРНК) і навіть деякі молекули ДНК не є генетичними, бо не утворюють генів. Однак вони забезпечують реалізацію генетичної інформацію в клітині, тобто синтез закодованих у генах поліпептидів.
У природі роль гНК виконують як ДНК у більшості видів організмів, так і РНК (віруси).
В клітинах еукаріотів генетична ДНК дволанцюгова і входить до складу надмолекулярних комплексів - хромосом, мітохондрій, пластид.
Молекули генетичної ДНК і РНК можуть бути лінійними і кільцевими одноланцюговими і дволанцюговими. Інформація з ДНК переписується на різні за будовою РНК, молекули яких разом із рибосомами забезпечують синтез відповідних поліпептидів.
За структуризацією нуклеїнові кислоти поділяються на декілька
рівнів.
Первинна структура. ДНК і РНК являють собою біополімер, утворений нуклеотидами, що повторюються. Кожний нуклеотид складається із трьох компонентів:
1. Циклічна сполука, що містить азот і зветься азотною основою. Відомо 5 азотних основ: аденін (А), тимін (Т), гуанін (Г), цитозін (Ц), урацил (У). Урацил виявлений тільки у РНК, тимін — ДНК. Двоциклічні основи - аденін і гуанін - належать до складу пуринів, моноциклічні - цитозин, тимін і урацил - до піримідинів.
2. Цукор - пентоза, що об’єднує 5 атомів вуглецю. До складу ДНК входить - дезоксирибоза, до складу РНК - рибоза.
3. Фосфорна кислота.
Одна із важливих особливостей ДНК описується правилом еквівалентності, сутність якого полягає в тому, що в молекулах ДНК і РНК молярні відношення пуринів до піримідинів дорівнює одиниці.
![]()
![]()
Ці правила виявили стійкий взаємозв’язок вмісту А і Т, Ц і Г. Ця закономірність, відкрита Е. Чаргафом в 1950 році (українець за походженням, який працював у США), відіграла важливу роль у створенні дволанцюгової моделі ДНК.
Вуглеводний компонент у складі нуклеотиду з’єднується з гетероциклічною основою N - глікозидним зв’язком.
Залишки нуклеотидів мономерів у нуклеїнових кислотах з’єднані між собою фосфодиефірними зв’язками, які відбуваються між 3' - вуглецевим атомом одного нуклеотидного залишку і 5' - атомом іншого. Тому зв’язок між двома сусідніми нуклеотидами називають 3' - 5' - фосфодиефірними.
Полінуклеотидні ланцюги ДНК і РНК-полярні: на одному кінці іавжди знаходиться вільна або заміщена група 3'-ОН, а на протилежному 5'- ОН.
Таким чином, до первинної структури ДНК і РНК входить будова їх мономерних залишків, хімічна природа міжнуклеотидих ковалентних зв’язків і послідовність розташування мономерних ланок у полінуклеотидному ланцюгу.
Просторова організація ДНК і РНК визначається їх нуклеотидними послідовностями через структури двох рівнів - вторинну і третинну.
Вторинна структура — (певний ступінь спіральності) створюється взаємодією в полінуклеотидному ланцюгу сусідніх нуклеотидів, а у випадку двоспіральних молекул також взаємодією нуклеотидних залишків, що знаходяться один проти одного у подвійній спіралі.
В 1953 році Дж. Уотсон і Ф. Крік збудували першу модель молекулярної організації ДНК, згідно з якою ця молекула являє собою праву спіраль, утворену двома полінуклеотидним ланцюгами, закрученими один на одному і навколо спільної осі. Автори довели, що це правило виконується в тому випадку, якщо аденін (А) одного ланцюга утворює стабілізовану водневими зв’язками пару з тиміном (Т), а гуанін (Г) - з цитозином (Ц). Ці взаємодіючі пари отримали назву комплементарних.
Свою модель ДНК Уотсон і Крік запропонували, спираючись на дані рентгеноструктурного аналізу М.Уілкінса і Р.Франкліна і правила Чаргоффа.
ДНК - це дволанцюгова спіральна структура, в середині якої знаходяться спарені азотні основи (А - Т) (Г - Ц), а зовні - фосфатні групи нуклеотидів.
Спіраль закручена таким чином, що на її поверхні утворюється 2 борозни: велика (шириною біля 2,20 нм) і мала 1,2 нм. Полінуклеотидні ланцюги у спіралі ДНК антипаралельні, Тобто на кожному з кінців лінійної дволанцюгової молекули ДНК розташовані 5' - кінець одного і 3' - кінець другого ланцюга. Діаметр спіралі дорівнює 1,8 нм, довжина витка приблизно 3,4 нм, в одному витку спіралі вміщується 10 нуклеотидних залишків (Мал. 2.5).
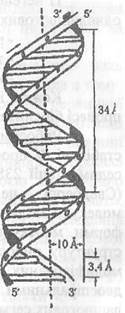
Малюнок 2.5
Модель двониткової спіральної молекули ДНК
Пізніше було з’ясовано, що модель Уотсона і Кріка описує структуру однієї найбільш розповсюдженої подвійної спіралі, що була названа В - формою, або В - конформацією. Існують і інші форми ДНК, які можуть взаємно переходити одна в одну(А,С, Z).
Третинна структура ДНК. У природі існують лінійні й кільцеві молекули ДНК. Лінійна молекула в ядрах еукаріотів упакована в компактну структуру і займає всього 1/5 об’єму клітини. Довжина ДНК хромосомі людини може досягти 8 см, але компактизована вона так, що вміщується у хромосомі довжиною 5 нм. Отже, спіралізована молекула ДНК у подальшому може утворювати більш компактні структури, в тому числі кільцеву форму і суперспіраль.
НадспІралізація ДНК властива хромосомам еукаріотів, а також вірусів і бактерій.
Молекули ДНК мітохондрій, хлоропластів, хромосом і плазмідій бактерій, а також ДНК багатьох вірусів мають кільцеву форму, майже вільну від білків і надспіралізовані.
Крім подвійних спіралей, зустрічаються і одноланцюгові ДНК, ДНК фагів фХ174, М13 та інші.
Макромолекулярна структура РНК. Переважна більшість РНК живих істот є одноланцюговими. Однак, у деяких вірусів (реовірус та ін.) зустрічаються дволанцюгові РНК, які виконують роль носіїв генетичної інформації. Крім того, дволанцюгові РНК утворюються в процесі розмноження деяких вірусів, у яких генетична РНК є однолащдоговою. Геномна РНК майже у всіх вірусів лінійна (вірус тютюнової мозаїки, поліомеліту, грипу).
Із негенетичних РНК у клітинах еукаріотів існує гри основних типи одноланцюгових молекул:
- рибосомні РНК (рРНК),
- транспортні РНК (тРНК),
- матричні або інформаційні РНК (іРНК), (мРНК).
Кожен із цих типів виконує свою специфічну роль у складному процесі біосинтезу білка.
Рибосомні РНК складають до 90% усієї РНК клітини, вони досить стабільні. У прокаріотів розрізняють три різних типи рРНК із коефіцієнтом седиментації 23S, 16S, 5S; у еукаріотів - чотири: 28S, 18S, 5S, 58S. S - (Сведбсрг) - це одиниця коефіцієнту, який є пропорційним седиментації молекули за даного відцентрового прискорення і, отже, залежить від маси і форми молекули. Всі рРНК разом із рибосомними білками входять у структуру рибосом. Для первинної структури рРНК характерна наявність модифікованих основ. Вторинна структура представлена короткими двоспіральними ділянками, які утворюються поєднанням зігнутих одно; ланцюгових сегментів однієї і тієї ж молекули.
Транспортні РНК являють собою одноланцюгові молекули, що складаються із 70 - 90 нукпеотидів, константа седиментації - 4S. У клітині на долю тРНК припадає 10-20%. Молекула тРНК здатна ковалентно зв’язуватися з відповідною їх амінокислотою і приєднуватися через систему водневих зв’язків до одного із триплетів (кодонів) молекули іРНК. Таким чином тРНК реалізують кодову відповідність між колонами іРНК і амінокислотами. Третинна структура дуже компактна, так що вся молекула має вигляд палички, загнутої у вигляді букви Г. У більшості клітин еукаріотів у тримується біля 60 тРНК. Це означає, що для кожної амінокислоти існує більш ніж по одній тРНК. Різні молекули тРНК, що акцентують одну і ту ж амінокислоту, називаються ізоакцепторами.
Інформаційна (матрична) РНК (іРНК) (мРНК) містить генетичну інформацію про послідовність амінокислот у поліпептидах і слугує матрицею її ні їх біосинтезу. Первинна структура іРНК містить кодони, відповідні кодонам ДНК, на матриці якої ця іРНК синтезується. Маса іРНК у клітині складає не більше 5% загальної кількості РНК. Окремі молекули іРНК гетерогенні, за розмірами константи седиментації цих молекул коливаються під декількох одиниць Сведберга до 20S і більше залежно від того, якої протяжності поліпептид кодується. мРНК дуже активна метаболічно у порівнянні з іншими типами РНК і являє собою дуже лабільну, короткоживучу форму. В еукаріотів ІРНК більш стабільні; їх існування в клітині може вимірюватися десятками хвилин, годинами і навіть днями.