ОСНОВИ МЕДИЧНОЇ БІОЛОГІЇ - 2012
Організація потоку інформації у клітині. Регуляція експресії генів. Молекулярні механізми мінливості в людини
Білки визначають структуру, форму, і функції клітини. На їх частку припадає майже половина сухої речовини. Проте в природі не доведено жодного випадку, коли б молекула білка (поліпептиду) синтезувала собі подібну. Біологічна інформація зберігається в генах у структурі ДНК у такій формі, що може копіюватися і передаватися наступним клітинам. Передача інформації розпочинається з реплікації (поділ ДНК) з утворенням двох ланцюгів ДНК. Під час транскрипції відбувається перенесення генетичної інформації з ДНК на РНК - це перший крок потоку біологічної інформації. На ступний крок - трансляція - перенесення інформації з РНК до білка.
Таким чином, генетична інформація записана в лінійній послідовності нуклеотидів ДНК (в ядрі клітини). За участі РНК ця інформація надходить (транслюється) до рибосом у цитоплазму з утворенням поліпептиду з амінокислот. Потік інформації відбувається таким шляхом:
ДНК(ген) — ► про-іРНК— ► іРНК—► поліпептид - ►білок.
Біосинтез білка, його основні етап. Біосинтез білка - це багатоступінчастий ферментативний процес, який здійснюється в клітинах живих організмів і забезпечує постійне оновлення білків органів і тканин. Процес синтезу білка складається з чотирьох послідовних стадій, або етапів: транскрипція, процесінг (постранскрипційні процеси), трансляція,
посттрансляційна модифікація (процесінг білку).
Транскрипція (від лат. transcription - переписування) - це складний біологічний процес реакції матричного синтезу, який забезпечує зчитування спадкової інформації з послідовності нуклеотидів ДНК на послідовність нуклеотидів РНК. Транскрипція — процес ферментативний, енергозалежний. Відбувається він під час інтерфази. На ДНК в результаті транскрипції утворюються всі види РНК: іРНК, тРНК, рРНК. Білки пістони, що забезпечують структурну організацію хромосом, також є репресорами в процесі зчитування генетичної інформації. Тому початок переписування генетичної інформації пов’язаний зі звільнення певної ділянки ланцюга ДНК (гена) від гістонів за допомогою негістонових білків. Вони мають здатність розпізнавати гени, ген активується, з нього зчитується генетична інформація, і цим забезпечується синтез необхідних білків. Транскрипція каталізується ферментами ДНК-залежними РНК-полімеразами (РНК-полімеразами). У прокаріотів існує один тип РНК-полімерази, який каталізує синтез всіх класів РНК. В еукаріотів існують три типи РНК-полімераз (І, II і III): І - каталізує синтез рРНК, II - синтез про-іРНК, III - синтез тРНК. У мітохондріях існує своя власна РНК-полімераза. Ділянка ДНК між промотором і термінатором, яка транскрибується, називається одиницею транскрипції. Молекула РНК, яка при цьому утворюється, називається первинним транскриптом або про-іРНК.
Процес транскрипції включає три етапи:
1. Ініціація.
2. Елонгація.
3. Термінація.
Ініціація. Певна ділянка молекули ДНК за сигналом з цитоплазми починає розкручуватись і розділятися на два ланцюги. В цьому процесі приймають участь фермент хеліказа (геліказа), який роз’єднує комплементарні водневі зв’язки між азотистими основами та дестабілізуючі SSB-білки, які запобігають самозбірці ДНК. Синтез і-РНК починається із знаходження РНК-полімеразою ділянки ДНК, яка вказує на початок транскрипції — стартовий кодон промотора гену. РНК-полімераза приєднується тільки до одного кодогенного (кодуючого) ланцюга ДНК в певній ділянці — початку гену (внаслідок того, що РНК-полімераза здатна приєднувати нуклеотиди від 5'-кінця ланцюга до 3'-кінця, і тільки один ланцюг починається з 3-кінця (3'-5'), його й називають кодогенним). Ініціація завершується з’єднанням двох перших рибонуклеозидтрифосфатів.
Елонгація — нарощування ланцюга РНК. Рухаючись вздовж, РНК-полімераза забезпечує точне зчитування та приєднання рибонуклеозидтрифосфатів: АТФ, ГФФ, ЦТФ, УТФ, комплементарних матриці (кодогенному ланцюгу ДНК).
Термінація. Елонгація продовжується до того часу, поки РНК-полімераза не доходить до послідовності — термінатора транскрипції (стоп-кодона або стоп-сигнала: АТТ, АЦТ, АТЦ). Синтез РНК припиняється, РНК-полімераза і синтезована про-іРНК звільняються з гену.
Під час транскрипції ДНК поступово, після проходження РНК-полімерази, знову з’єднується у подвійну спіраль.
Ділянка молекули ДНК, що містить промотор, послідовність, яка транскрибується, та термінатор, називається транскриптоном. Утворена під час транскрипції РНК (РНК- попередник) є точною комплементарною копією ділянки ДНК транскриптону (гену) включаючи ділянки, які не визначають послідовність амінокислот, тому в ядрі відбуваються процеси «дозрівання» РНК та утворення РНК (і-РНК, т-РНК, р-РНК), які будуть виконувати свою функцію під час синтезу білків на рибосомах.
Процесинг — «дозрівання» різних типів РНК - утворення з РНК-попередника (про-іРНК) зрілої функціонуючої і-РНК. У клітинах прокаріотів іРНК синтезується відразу в зрілому вигляді і готова до виконання своєї біологічної функції. В еукаріотів спочатку утворюється незріла іРНК (первинний транскрипт, іРНК-попередниця або про-іРНК, ядерна гетерогенна РНК), яка зазнає посттранскрипційної модифікації - процесингу. Процесинг включає наступні процеси: 1) приєднання до 5'-кінця молекули про-іРНК «шапочки», або «кепа» (англ. cap - шапочка). «Keп» являє собою метилгуанозин. Вважають, що він захищає іРНК від руйнування клітинними нуклеазами і забезпечує розпізнавання її малої субодиниці рибосомами; 2) приєднання послідовності нуклеотидів, які мають функціональне значення: лідер — забезпечує приєднання і-РНК до малої субодиниці рибосоми, стартовий кодон — триплет нуклеотидів (АУГ), кодуючий амінокислоту метіонін; трейлер (послідовність нуклеотидів, яка включає нонсенс-кодон); 3) приєднання до 3'-кінця молекули про-іРНК послідовності з 20-30 аденілових нуклеотидів ("хвоста"), яка надає молекулі стабільності; 4) структурні гени (ДНК) еукаріотів містять кодуючі ділянки - екзони (кодують амінокислоти) і некодучі ділянки - інтрони (не кодують амінокислот). Оскільки про-іРНК є комплементарною копією гена (ДНК), вона теж містить екзони і інтрони. Під час процесинга інтрони за допомогою ферментів рестриктаз вирізаються з молекули про-іРНК, а екзони за допомогою ферментів лігаз зшиваються з утворенням зрілої іРНК. Цей процес називається сплайсингом. Зріла іРНК, зрозуміло, менша за розмірами від про-іРНК. Зріла іРНК з «кепом» на 5'-кінці і поліаденіловою послідовністю (А-А-А-А-) на 3'-кінці через пори в ядерній оболонці виходить у цитоплазму (часто у вигляді комплексів з білками - інформосом). Білкові молекули захищають іРНК від руйнування. Функціонувати вона починає тільки після звільнення від білка.
Трансляція — складний біологічний процес реакції матричного синтезу, який забезпечує передачу спадкової інформації з послідовності нуклеотидів іРНК на послідовність амінокислот поліпептиду. Трансляція — ферментативний, енергозалежний процес, який в еукаріотичній клітині відбувається в цитоплазмі на рибосомах.
Трансляція включає три фази: ініціації, елонгації, термінації.
Ініціації — процес об’єднання іРНК з субодиницями рибосом та тРНК. Розпочинається цей процес з активації амінокислот, які вступають в реакцію з АТФ. Реакцію каталізує фермент аміноацил-тРНК-синтетаза, який для кожної амінокислоти свій. Активована амінокислота приєднується до своєї специфічної тРНК. Утворений комплекс називають навантаженою тРНК, а процес розпізнавання амінокислот тРНК, називають рекогніцією.
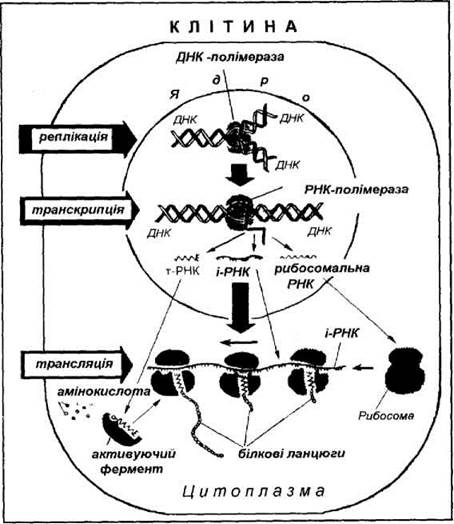
Рис. 10. Схема біосинтезу білка
Процес дозрівання проходять також тРНК і рРНК.
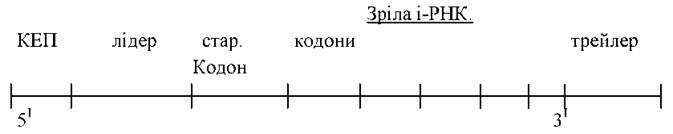
Ланцюг іРНК об’єднується з малою субодиницею рибосоми шляхом утворення водневих зв’язків між комплементарними парами відповідних азотистих основ іРНК та рРНК. Амінокислота метіонін ініціює процес синтезу. До стартового кодону АУГ приєднується аміноаціл-тРНК (з антикодоном УАЦ), яка транспортує метіонін. Утворюється комплекс ініціації. Потім до комплексу приєднується велика субодиниця рибосоми. Утворюється активна рибосома, яка має два активних центри: аміноацильний (А) та пептидильний (П). Процес ініціації відбувається за участю факторів ініціації — лабільних білків, які зв’язуються з малими субодиницями рибосом і які по завершенню ініціації відділяються від рибосом.
В цитоплазмі постійно відбувається утворення сполук тРНК з амінокислотами. тРНК специфічно з’єднуються з амінокислотами за допомогою ферментів аміноацил — т-РНК — синтетаз, що визначають відповідну тРНК та приєднують до неї амінокислоту.
Елонгація — нарощування поліпептидного ланцюга, це циклічні приєднання амінокислот з моменту утворення першого пептидного зв’язку до останнього.
Перша т-РНК звільняється від кодону і-РНК та залишає П-центр, утворений дипептид знаходиться в А-центрі з’єднаний з другою т-РНК. Між амінокислотами утворюється пептидний зв’язок, який каталізується білками-ферментами, що входять до складу рибосоми.
Далі відбувається рух системи на один крок-кодон. Друга т-РНК з дипептидом надходить в П-центр, А-центр звільняється, слідуюча тРНК з проліном вступає в контакт з А-центром. До третього кодону приєднується специфічна аміноацил-т-РНК з третьою амінокислотою. Між дипептидом та третьою амінокислотою утворюється пептидний зв’язок. Друга т-РНК звільнюється... і так крок за кроком відбувається нарощування поліпептидного ланцюга поки з А-центром не зв’язується термінальний кодон для якого не існує специфічної комплементарної т-РНК.
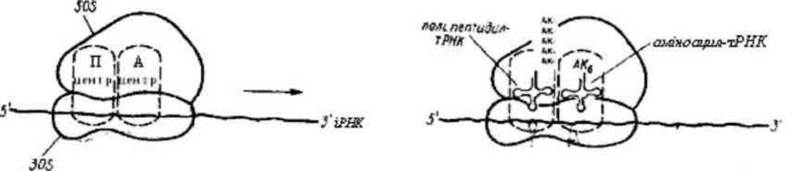
Рис. 11. Елонгація поліпептидного ланцюга: АК-АК-амінокислоти
Термінація синтезу поліпептидного ланцюга (завершення синтезу поліпептида) пов’язана із надходженням в А-центрі термінального кодону (наприклад, АГЦ). До термінального стоп-кодону приєднується специфічний рибосомальний білок. До останньої амінокислоти приєднується вода і поліпептид відділяється від тРНК, внаслідок пептидний ланцюг втрачає зв’язки з рибосомою, яка розкладається на 2 субодиниці.
Фаза елонгації у синтезу білку
1-й етап - аміноацил-тРНК приєднується до кодону, який розміщений в А-центрі;
2-й етап - між амінокислотами, які розміщені в А- та П- центрах, утворюється пептидний зв’язок; тРНК, яка розміщена в П-центрі, звільняється від своєї амінокислоти і покидає рибосому;
3-й етап - рибосома рухається по іРНК, робить крок на один кодон так, що тРНК з пептидним ланцюжком переходить з А-центра в П-центр; вільний А-центр рибосоми може бути зайнятий тільки відповідною аміноацил-тРНК.
Термінація синтезу поліпептидного ланцюга
1-й етап - приєднання фактора звільнення до стоп-кодону;
2-й етап - термінація звільнення завершеного пептиду;
3-й етап - дисоціація рибосоми на дві субодиниці.
Посттрансляційні перетворення білків
У рибосомі в процесі трансляції синтезуються поліпептидні ланцюги білкової молекули, що відповідають первинній структурі білка, яка не має метаболічної активності. Далі синтезовані поліпептиди надходять у цитоплазму, ендоплазматичну сітку або комплекс Гольджи, де формуються вторинні, третинні та четвертині структури білка. Синтезовані молекули об’єднуються з вуглеводним и або ліпідними молекулами. Залежно від того, які функції білка (фермент, антитіло, будівельний і т.д.), він бере участь у забезпеченні морфофункціональних особливостей клітини (організму), тобто в формуванні окремих складних ознак. Це кінцевий етап процесу реалізації генетичної інформації.
Екзонно-інтронна організація генів еукаріот
У еукаріотів гени принципово однаково організовані з генами прокаріот, але мають деякі особливості. Відмінності полягають в самій організації генетичного матеріалу. У прокаріотів кодуючи гени є неперервними, тоді як у еукаріотів встановлена переривчаста будова генів. Структурні гени включають послідовності ДНК, які не визначають послідовності амінокислот кодуючих білків — їх називають інтрони. Послідовності, які включаються до складу зрілих і-РНК та визначають первинну структуру білків називають екзонами. Причому в структурних генах кодуючі ділянки — екзони, розділяються некодуючими ділянками — інтронами (мозаїчна будова гена). Вважається, що початок інтронів комплементарний його кінцю, тому після утворення РНК-попередника (гетероядерної РНК), під час процесінгу, інтрони утворюють петлі, які вирізаються ферментами нуклеазами, а екзони послідовно зшиваються (сплайсинг) РНК-лігазами. Утворюються зрілі і-РНК, які через ядерну оболонку надходять у цитоплазму і забезпечують процес трансляції (синтезу поліпептидного ланцюга). Таким чином, екзони — інформативні ділянки структурних генів, які визначають (кодують) амінокислотні послідовності - поліпептиди (білки). Інтрони — неінформативні ділянки структурних генів, які не визначають послідовності амінокислот. Кількість нуклеотидів в інтронних локусах в 5-10 разів більша, ніж в екзонах. Така організація структурних генів типова для еукаріот, у прокаріот інтронні ділянки не знайдені.
Присутність в еукаріотів чисельних інтронів полегшує генетичну рекомбінацію між екзонами і забезпечує більшу гнучкість у синтезі білка. Також можна вважати, що інтрони несуть запас інформації, який зумовлює мінливість. Доведено, що з однієї первинної РНК у різних тканинах утворюється не один, а декілька різних за довжиною іРНК-транскриптів. Виникнення нових білків забезпечує ефективність еволюції організмів.
Центральна догма молекулярної біології
Положення про те, що генетична інформація зберігається в ДНК і в такому вигляді в результаті реплікації передається від клітини до клітини в ряді поколінь і що вона реалізується в процесі біосинтеза білка шляхом передачі від ДНК до іРНК (транскрипція) і далі від іРНК до білка (трансляція), відоме як центральна догма молекулярної біології. Її запропонував у 1958 р. Ф. Крік. Вона може бути виражена у вигляді схеми:
![]()
Американські генетики Г. Темін і Д. Балтимор (1970) виявили в РНК-вмісних онкогенних вірусів фермент, який каталізує синтез ДНК на матриці іРНК (зворотна транскрипція). Цей фермент було названо РНК-залежна ДНК-полімераза, або зворотна транскриптаза - ревертаза (лат. reversio - повернення). У зв'язку з відкриттям зворотної транскрипції схема центральної догми молекулярної біології набула наступного вигляду:
![]()
Регуляція експресії генів у прокаріотів і еукаріотів
Усі соматичні клітини організму містять однаковий набір хромосом, однаковий набір генів і несуть повну генетичну інформацію, властиву даному виду. Проте клітини в одного і того самого організму різні за будовою, функцією, складом білків-ферментів. Основу цих відмінностей складає експресія (активність) різних частин генома. Генна експресія - це молекулярний механізм реалізації спадкової інформації, завдяки якому ген виявляє свій потенціал конкретною генотипною ознакою організму. У геномі існують дві групи генів: 1) конститутивні гени - експресуються постійно (рибосомні білки, РНК-полімерази, молекули тРНК, рРНК, ін.); 2) неконституційні — неактивні, репресовані, але при необхідності можуть активуватися клітинними, тканинними регуляторами або гормонами. Вони діють на певних фазах клітинного циклу, забезпечують цитодиференціювання, специфічність функціонування клітин в багатоклітинному організмі, пристосованість в умовах середовища
Механізм регуляції експресії генів вперше в 1961 р. пояснили французькі мікробіологи Ф. Жакоб і Ж. Моно, які вивчали регуляцію синтезу ферментів у бактерії кишкової палички (Е.соїі). Цей механізм відомий як гіпотеза оперона (Нобелівська премія, 1968).
Оперон - генетична одиниця регуляції транскрипції генів у прокаріотів. Це послідовність нуклеотидів ДНК, яка об'єднує структурні гени, промотор, оператор і термінатор. Структурні гени тісно зчеплені між собою, розташовані один біля одного і несуть інформацію для синтезу ферментів послідовних реакцій одного метаболічного циклу. Функціонують структурні гени одного оперону як одне ціле. До оперона належать ділянки: промотор, оператор, структурні гени і термінатор. Промотор -ділянка ДНК оперона, до якої первинно прикріплюється РНК-полімераза і з якої починається транскрипція структурних генів. Оператор - ділянка ДНК оперона, яка розташована між промотором і структурними генами і з якою може зв'язуватися білок-репресор. Структурні гени синтезують матричну РНК, коли оператор включений, і припиняють синтез, коли оператор виключений. Оператор включається або виключається білком-репресором. Синтез білка-репресора кодується геном-регулятором, який може розміщуватися поруч з опероном або на відстані від нього. Білок-репресор може бути в двох формах - активній і неактивній. Перебуваючи в активній формі, репресор зв'язується з оператором і виключає його. Це виключає із функції весь оперон (транскрипція не відбувається). Включення оперона відбувається в тому випадку, коли білок-репресор знаходиться в неактивній формі і не здатний зв'язатися з оператором. У регуляції експресії генів беруть участь негенетичні фактори - ефектори. Ефектори — низькомолекулярні речовини, які взаємодіють з білками-репресорами і змінюють їх здатність зв'язуватися з оператором. Якщо комплекс (оператор+білок-регулятор) виключає транскрипцію, то такий контроль називається негативним, якщо включає - позитивним.
Класичним прикладом роботи оперона за типом індукції є модель лактозного оперона (lас-оперона) бактерії кишкової палички. Lac-оперон містить три структурних гена, які кодують синтез трьох ферментів, необхідних для метаболізму лактози. Робота оперона знаходиться під контролем гена-регулятора, який кодує синтез активного білка-репресора. За умови відсутності лактози білок-репресор зв'язується з оператором, транскрипція не відбувається (негативний контроль). Якщо в культуральному середовищі з'являється лактоза (замість глюкози), вона проникає в клітину і зв'язується з білком-репресором, він стає неактивним і не може зв'язатися з оператором. Оператор звільняється від білка-репресора і структурні гени можуть працювати (транскрипція відбувається). Поки в середовищі знаходиться лактоза, до тих пір працює оперон і утворюються ферменти, необхідні для її розщеплення. Таким чином, лактоза виступає одночасно як субстрат розщеплення і як індуктор свого власного розщеплення. Негативний контроль роботи lac-оперона пояснює сутність феномену індукції: немає індуктора (лактози) - оперон не працює, є індуктор(лактоза) - оперон активно працює.
Синтез ферментів може не тільки індукуватися, але й репресуватися. Поки кінцевий продукт D синтезується в необхідній кількості, репресор перебуває в неактивному стані, ген-оператор «включений» і структурні гени працюють. Як тільки продукт D накопичується в кількостях більших, ніж це потрібно клітині, він вступає в реакцію з репресором, останній активується і, вступаючи у взаємодію з геном-оператором, припиняє роботу всієї системи и синтез ферментів зупиняється.
Таким чином, вихідна речовина, яка надійшла у клітину (субстрат реакції), є індуктором, що запускає роботу оперона; гальмування синтезу відбувається кінцевим продуктом, який утворюється в результаті реакції (принцип зворотного зв’язку).
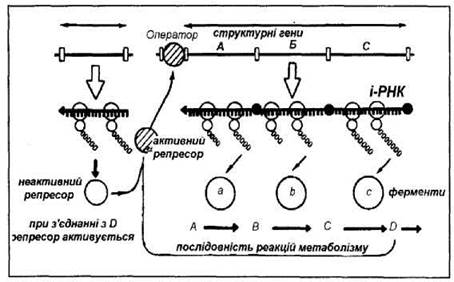
Рис. 12. Схема механізму генетичного контролю синтезу ферментів у бактерій (за: Жакоб і Моно, 1961)
Отже, генетична система клітини, використовуючи механізми індукції і репресії, сприймає сигнали щодо початку і закінчення синтезу того або іншого білка (ферменту) і здійснює цей процес з певною швидкістю.
Експресія генів еукаріотів має свої особливості (більш складний характер). За схемою Г.П. Георгієва (1972), кожен оперон складається з проксимальної акцепторної зони, яка включає ряд послідовно розташованих генів (операторів і промоторів), і дистальної (регуляторної зони). Акцепторна зона керує дією структурних генів через білки-репресори. Один структурний ген може багаторазово повторюватися, утворюючи цілу серію однакових послідовностей. Структурні гени, які відповідають за різні ланки одного ланцюга біохімічних реакцій, можуть бути зосереджені не в одному опероні, а розсіяні по геному.
У еукаріотів існує шлях регуляції, якого не має у прокаріотів - одночасне пригнічення активності генів у цілій хромосомі або у її ділянці. Такий шлях регуляції здійснюється білками- гістонами, що входять до складу хромосом.
Особливості експресії в еукаріотів полягають: 1) в еукаріотів просторово розділені процеси транскрипції (відбувається у ядрі) та трансляції (відбувається у цитоплазмі); 2) в еукаріотів наявна ампліфікація генів - багаторазове збільшення чисельності копій генів для інтенсивного синтезу молекул; 3) геноми вищих еукаріотів значно складніші (наприклад, у людини гаплоїдний набор включає 23 хромосоми - приблизно 40000 генів); 4) на відміну від прокаріотів, в еукаріотів гени ферментів певного метаболічного циклу можуть бути розташовані у різних хромосомах, тому синтезована іРНК є моноцистронною; 5) особливістю геному еукаріотів є наявність енхансерів - сегменти ДНК, які виконують роль «підсилювачів» в процесі зчитування генетичної інформації; 6) у багатоклітинному організмі на геном еукаріотів постійний вплив має ендокринна система; 7) на стадії трансляції у еукаріотів також здійснюється контроль експресії генів; 8) геном еукаріотів надлишковий: деякі гени багаторазово повторюються, а інші зовсім не відіграють генетичної ролі (сателітна ДНК); 9) регуляція експресії генів еукаріотів може здійснюватись га стадії пост трансляційних змін.