Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Вищі функції нервової системи умовні рефлекси, навчання та споріднені явища
Навчання та пам'ять
Характерна ознака живих організмів, передусім людини, - здатність змінювати поведінку на підставі набутого досвіду. Це відбувається завдяки навчанню, що є процесом отримання інформації, та пам’яті, тобто зберігання та нагромадження інформації. Названі два тісно споріднені явища треба розглядати нерозривно одне від одного.
Форми пам’яті
З фізіологічного погляду пам’ять поділяють на свідому й підсвідому (табл. 16-1). Свідома пам’ять, яку також називають декларативною, чи пізнавальною, пов’язана зі станом свідомості чи, принаймні, неспання. За її зберігання відповідає морський коник та інші ділянки медіальних скроневих часток мозку. Цю форму пам’яті поділяють так: на події (епізодична пам’ять) та пам’ять на слова, правила, мову тощо (семантична пам’ять). Підсвідома пам’ять не стосується стану неспання, її також називають недекларативною, чи зворотною, а зберігання її в більшості випадків не пов’язане з морським коником. До підсвідомої пам’яті належать уміння, навички, умовні рефлекси та ін. (див. табл. 16-1). Щоправда, свідома пам’ять, необхідна, скажімо, для їзди на велосипеді, стає підсвідомою, як тільки людина навчиться добре це робити.
Свідома та багато форм підсвідомої пам’яті пов’язані, по-перше, із короткочасною пам’яттю, що триває від кількох секунд до кількох годин, упродовж яких процеси в морському конику та інших ділянках закладають довгочасні зміни у силі синаптичних зв’язків; та, по-друге, із довготривалою пам’яттю, яка зберігає спогади роками, а інколи - усе життя. Спогади, які зберігає короткочасна пам’ять, піддаються руйнуванню внаслідок травм чи дії різноманітних ліків, натомість спогади довготривалої пам’яті стійкіші. Робоча пам’ять - це форма короткочасної пам’яті, яка зберігає інформацію доступною, як звичайно, протягом дуже коротких періодів часу, коли особа планує на її підставі якусь дію.
Підсвідома пам’ять
Як видно з табл. 16-1, підсвідома пам’ять охоплює вміння й навички, що одразу ж після набуття стають несвідомими та автоматичними, а також натаскування, тобто полегшення розпізнавання слів чи об’єктів за допомогою їхньої попередньої демонстрації. Як приклад можна навести ситуацію, за якої особі легше пригадати слово, якщо їй підказують кілька перших букв.

Рис. 16-1. Отримане за допомогою ПЕТ сканування зображення лівої півкулі головного мозку. Показано зони найвищої нейронної активності, коли суб’єкт виконує різні дії. А - дивиться на слова; активна первинна зорова кора та частина зорово-асоціативної кори. В - слухає слова; активна зона на стику скроневої та тім’яної ділянок кори. С - вимовляє слова; активна зона Брока та прилегла лобова частка. D - думає про слова; активні великі зони, в тому числі значна частина лобової частки (на підставі ПЕТ сканування з Kandel ER, Schwartz JH, Jesseli TM: Essentials of Neural Science and Behavior. McGraw-Hill, 1995).
Інші форми підсвідомої пам’яті можна розділити на неасоціативні та асоціативні. У разі неасоціативного навчання організм вивчає окремий подразник, у разі асоціативного навчання відбувається вивчення взаємозв’язків між різними подразниками.
Набування навичок та сенсибілізація
Набування навичок - це проста форма навчання, за якої нейтральний подразник часто повторюють. За першим разом цей подразник є повним і зумовлює реакцію (орієнтувальний рефлекс, або реакцію «що це?»); зі збільшенням кількості повторювань він спричинює щораз слабшу біоелектричну реакцію. Зрештою, суб’єкт звикає до подразника та ігнорує його. Сенсибілізація є в певному сенсі протилежною. Повторюваний подразник спричинює сильнішу реакцію, якщо його поєднують один чи кілька разів із неприємним або приємним подразником. Загальновідомо, що у людей від подразника виникає інтенсифікація рівня збудження. Наприклад, мама, сну якої не заважають різноманітні шуми, моментально прокидається від плачу своєї дитини.
Процес набування навичок є класичним прикладом неасоціативного навчання. Класичний приклад асоціативного навчання - умовний рефлекс.
Таблиця 16-1. Типи пам’яті

Умовні рефлекси
Умовний (набутий) рефлекс є рефлекторною реакцією на подразник, який раніше спричиняв слабку реакцію, набуту шляхом повторюваного поєднання цього подразника з іншим, який подібну реакцію зумовлює, або ж не спричиняв жодної. У класичному експерименті вироблення умовного рефлексу Павлова вивчали слиновиділення, зумовлюване вкладанням у рот собаки шматка м’яса. Безпосередньо перед цим дзвонили у дзвоник, повторюючи це доти, доки у собаки не почалося слиновиділення на звук дзвоника, навіть якщо їй не давали м’яса. У цьому експерименті м’ясо, яке вкладали в рот собаки, відігравало роль безумовного подразника (БП), тобто подразника, що, як звичайно, спричинює конкретну природну реакцію. Умовним подразником (УП) був звук дзвоника. Після поєднання безумовного та умовного подразників необхідну кількість разів УП зумовив реакцію, яку первинно спричиняв лише БП. Для цього потрібно, щоб УП передував БП. Інший приклад показано на рис. 16-2. Це так зване класичне вироблення умовного рефлексу. Сьогодні можна зробити так, щоб багато соматичних, вісцеральних та інших невральних змін, які відбуваються, стали умовними рефлекторними реакціями. Вироблення цих реакцій часто називають біологічним зворотним зв’язком. Зміни, які в цьому випадку можуть виникати, охоплюють альтерацію частоти серцевих скорочень та кров’яного тиску. Вважають, що за допомогою зумовлених знижень кров’яного тиску можна лікувати гіпертензію. Щоправда, депресорні реакції, спричинені у такий спосіб, є незначними.
Якщо У П діє тривалий час без БП, то умовний рефлекс, зрештою, зникає. Цей процес називають загасанням, чи внутрішнім гальмуванням. Якщо ж на тварину відразу після УП вплине зовнішній подразник, то умовної реакції може не відбутися (зовнішнє гальмування). Однак якщо умовний рефлекс підсилювати час від часу поєднанням УП та БП, то він не зникне.
Як зазначено в Розділі 15, умовні рефлекси важко формувати, якщо БП не поєднаний з приємним чи неприємним впливами. Сильним БП є подразнення як системи винагороди мозку (приємне, чи позитивне підсилення), так і системи уникання, або больовий шок шкіри (неприємне, чи негативне, підсилення). Оперантне (інструментальне) вироблення умовного рефлексу - це така форма вироблення рефлексів, за якої тварину вчать виконувати певні завдання для того, щоб вона отримала винагороду чи уникла покарання. Як БП у цьому разі є приємна чи неприємна подія, а УП - світло чи інший сигнал, що спонукає тварину виконувати завдання. Зумовлені рухові реакції, що дають змогу тварині уникнути неприємної події, називають умовними рефлексами уникнення. Наприклад, тварину вчать, що, натискаючи на планку, вона зможе уникнути удару електричним струMOM по кінцівках. Іншим прикладом є умовний рефлекс відрази до їжі. У тварин виникає сильна відраза до певної категорії їжі, якщо під час приймання цієї їжі вводити їй ін’єкцію препарату, який спричинює нудоту чи хворобу. Подібні реакції відрази трапляються у людей. Ці умовні реакції дуже сильні, інколи їх можна вивчати у випадку одноразового поєднання УП та БП і, на відміну від інших умовних рефлексів, вони розвиватимуться тоді, коли УП та БП розділені в часі (годину чи й більше). Вироблення умовних рефлексів відрази до їжі має важливе значення для виживання, оскільки ці рефлекси допомагають уникати отрут. Отже, не дивно, що мозок, можливо, генетично запрограмований на те, щоб сприяти розвитку реакцій відрази до їжі.
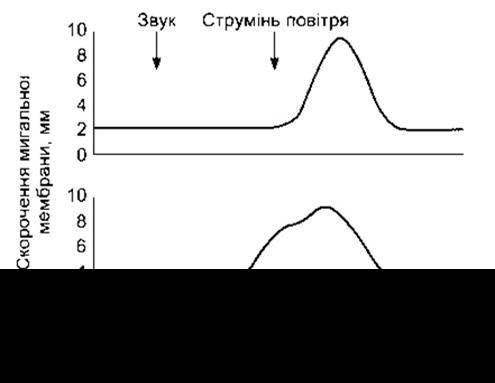
Рис. 16-2. Умовний рефлекс у кролика. Першою стрілкою позначено звук, який подають (УП), другою - ніжний струмінь повітря (БП), який спрямовують кроликові в око. Спочатку мигальна мембрана витягується, реагуючи тільки на струмінь повітря (вгорі). Після кількаразових повторювань двох подразників у парі мигальна мембрана починає скорочуватись, реагуючи на звук, що передує струменю повітря (внизу) (модифіковано з Alkon DL-Memory storage and neural systems. Sei Am [July] 1989;261:42).
Міжкіркове переміщення пам’яті
Якщо кіт чи мавпа виявляють умовні рефлекси на візуальний подразник, маючи одне око закрите, а після цього їх тестують, переносячи пов’язку на інше око, то тварина також відображає збереження умовної реакції. Це відбувається навіть у випадку перерізаного зорового перехрестя, коли візуальний сигнал із кожного ока йде винятково до іпсилатеральної ділянки кори. Якщо ж, окрім перерізаного зорового перехрестя, розділені передня та задня спайки, а також мозолисте тіло («тварина з роздвоєним мозком»), то переміщення пам’яті не виникає. Експерименти з частковим розділенням мозолистого тіла свідчать про те, що переміщення пам’яті відбувається в передній ділянці мозолистого тіла. Подібні результати простежувалися в людей, вроджених без мозолистого тіла, або в тих, кому його хірургічно видалили з метою запобігти епілептичним нападам. Це доводить таке: невральне кодування, потрібне для того, щоб «запам’ятовувати одним оком те, що побачило інше», було перенесене до протилежної ділянки кори через спайки. Є докази того, що подібне переміщення інформації відбувається через інші сенсорні шляхи.
Молекулярна основа пам’яті
Ключовим пунктом у пам’яті є альтерація сили відповідних синаптичних зв’язків. Майже у всіх найпростіших випадках така зміна зумовлює синтезування білка та активування генів. Це відбувається під час зміни короткочасної робочої пам’яті довготривалою. У тварин набуттю довготривалих завчених реакцій можна запобігти, якщо протягом п’яти хвилин після кожного навчального заняття піддавати їх анестезії, електрошоку, гіпотермії або ж дії препаратів, антитіл чи олігонуклеотидів, які блокують синтезування білків. Якщо ж такі заходи вживати через чотири години після навчальних занять, то вони жодного впливу на набуття реакцій уже не матимуть.
У людей подібним явищем є втрата пам’яті про події, що відбулися безпосередньо перед струсом головного мозку чи електрошокової терапії (ретроградна амнезія). Така амнезія простежується впродовж триваліших періодів часу, ніж у піддослідних тварин (інколи протягом багатьох днів), однак вона не впливає на віддаленіші в часі спогади.
Біохімічні процеси, що відбуваються під час набуття навичок та сенсибілізації у Aplysia та інших безхребетних, вивчені детально. Разом із процесами, що є в основі кількох
форм довготривалої потенціації (ДТП) та довготривалої депресії (ДТД), вони розглянуті у Розділі 4.
Набуття навичок зумовлене зниженням рівня Са2+ у сенсорних закінченнях, що опосередковують реагування на певний подразник. Відповідно, сенсибілізацію спричинює пролонгація потенціалу дії у цих закінченнях, що приводить до збільшення внутрішньоклітинного Са2+, який сприяє вивільненню нейротрансмітера в разі екзоцитозу.
Класичне вироблення умовних рефлексів також зафіксовано в Aplysia та ссавців в ізольованому спинному мозку У Aplysia БП діє пресинаптично на закінчення нейронів, активованих УП. Унаслідок цього в клітині з’являється вільний Са2+, що спричинює довготривалу зміну молекули аденілатциклази.
Отже, внаслідок активування цього ензиму умовним подразником утворюється більше цАМФ. Це, відповідно, закриває К+-канали та продовжує тривалість потенціалів дії за допомогою механізму, описаного у Розділі 4. Вирішальним моментом тут є взаємозв’язок з БП, що діє незадовго після УП.
У Aplysia простежуються морфологічні відповідники до навчання та пам’яті. Наприклад, 40% відповідних сенсорних закінчень у нормальному стані містять активні зони, тоді як у тварин із набутими навичками активні зони мають 10% сенсорних закінчень, а у сенсибілізованих - 65%. Довготривала пам’ять приводить до активування генів, що зумовлюють збільшення кількості синаптичних контактів.
Кодування підсвідомої пам’яті у ссавців
Без сумніву, молекулярні процеси, подібні до тих, що відбуваються в Aplysia, є в основі деяких аспектів підсвідомої пам’яті у ссавців. Щоправда, тут задіяні процеси, які стосуються різноманітних частин ЦНС. Деякі дослідники стверджують, що в цьому бере участь також смугасте тіло.
Відомо, що процес навчання переривається внаслідок ушкодження базальних ядер. Окрім цього, є докази участі у процесі й мозочка. Наприклад, вестибулоочний рефлекс, (ністагм) що підтримує зорову фіксацію під час рухів головою (див. Розділ 9), можна пристосувати до нової позиції очей. Однак ця здатність буде втрачена у випадку уражень жмутка мозочка. Вироблення умовного рефлексу кліпання очима, коли для цього на око спрямовують струмінь повітря як УП (див. рис. 16-2), не можливе за умов ушкоджень проміжного ядра. У цьому разі імпульси, зумовлені БП, діють через нижню оливу та волокна, спрямовані вгору до кори головного мозку, щоб змінити реагування клітини Пуркіньє на тонус, який з’являється через ядра мосту головного мозку та мохоподібні волокна. Модифікація перенесеної мохоподібними волокнами деполяризації клітини Пуркіньє опосередкована спрямованими вгору волокнами і також відповідає за зміни у ВОР та вивчених м’язових рухах.
Кодування свідомої пам’яті
Кодування свідомих спогадів стосується робочої пам’яті у лобових частках та специфічного опрацювання у морському конику.
Робоча пам’ять
Як уже зазначено, робоча пам’ять утримує інформацію, що надходить, доступною протягом короткого періоду часу, доки особа вирішує, що з нею робити. Це така форма пам’яті, що дає нам змогу, скажімо, відшукавши у записній книжці номер телефону, пам’ятати його стільки часу, скільки потрібно, щоб підняти трубку і набрати його. Вона складається з так званої центральної виконавчої системи, що розташована у передлобовій ділянці кори головного мозку, та двох «систем повторювання» - вербальної системи для зберігання вербальних спогадів та паралельної візуоспатичної системи для зберігання візуальних і просторових характеристик об’єктів. Інформацію у ці системи повторювання спрямовує виконавча система.
Морський коник та медіальна скронева частка
Ділянки робочої пам’яті сполучені з морським коником та закрутками морського коника медіальної скроневої ділянки кори, що примикають (див. рис. 15-1 та 16-3). У людей двостороннє руйнування вентрального морського коника, або ж хвороба Альцгеймера, та подібні процеси, що руйнують СА1-нейрони, призводять до значних дефектів короткочасної пам’яті. Такі ж наслідки спричинюють двосторонні ураження у мавп. Водночас подібні ушкодження в людей не впливають на робочу пам’ять та пам’ять про минуле. Окрім цього, як звичайно, неушкодженими є процеси підсвідомої пам’яті. В контексті свідомої пам’яті такі особи поводять себе адекватно доти, доки концентрують увагу на тому, що роблять. Однак як тільки їх відволікти від заняття навіть на короткий час, то вся пам’ять про те, що вони нещодавно робили, буде втрачена. Отже, такі люди здатні засвоювати нові знання та зберігати спогади про події, що відбулися до ушкодження, проте не можуть формувати нові довготривалі спогади. Морський коник тісно пов’язаний із закруткою морського коника у медіальній лобовій частці кори (див. рис. 12-3). Сьогодні процеси пам’яті вивчають не лише за допомогою фЯМР, а й вимірюванням збуджених потенціалів (тобто потенціалів пов’язаних із подією (ППП)) у хворих на епілепсію з імплантованими електродами. Коли ці особи згадують слова, то підвищена активність простежується у їхній лівій лобовій частці та у лівій закрутці морського коника, натомість, коли вони згадують образи чи сцени, то активність фіксують у правій лобовій частці та закрутках морського коника з обох боків.
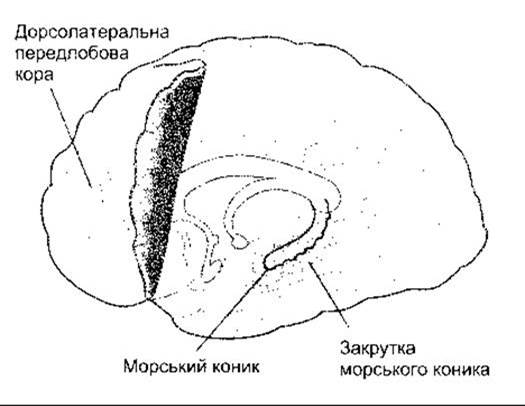
Рис. 16-3. Зони, задіяні у кодуванні свідомих спогадів (модифіковано з Russ MD: Memories are made of this. Science 1998; 281:1151).
У процесах пам’яті задіяні також зв’язки морського коника з проміжним мозком. У деяких алкоголіків із ушкодженням мозку погіршується пам’ять на недавні події. Таку втрату пам’яті пов’язують із наявністю патологічних змін у сосочкових тілах, що мають розгалужені еферентні з’єднання, які йдуть через склепіння до морського коника, і проекційні нервові волокна, які через сосочково-таламічний шлях проходять до переднього таламуса. Зазначимо, що ушкодження таламуса у мавп також спричинюють втрату пам’яті на недавні події. З таламуса волокна, що відповідають за пам’ять, відходять до передлобової ділянки кори, потім - до основи переднього мозку, звідки йдуть дифундивні холінергічні проекційні нервові волокна до всієї нової кори, мигдалеподібного тіла та морського коника з базального ядра Мейнерта. У випадку хвороби Альцгеймера простежуються серйозні втрати цих волокон (див. нижче).
Мигдалеподібне тіло тісно пов’язане з морським коником і задіяне у кодуванні емоційних спогадів. Як зазначено у Розділі 15, унаслідок ушкодження мигдалеподібних тіл тварини стають менш лякливими. Здорові люди запам’ятовують події, пов’язані з переживанням сильних емоцій, набагато ліпше, ніж події без емоційного навантаження. Натомість у пацієнтів із двосторонніми ушкодженнями мигдалеподібного тіла такої різниці нема.
Конфабуляція є цікавим, однак слабко вивченим станом, що інколи трапляється в осіб з ушкодженнями вентромедіальних ділянок лобових часток. Такі люди погано складають тести на пам’ять, однак невимушено описують події, яких ніколи не було. Такий стан ще називають «чесною брехнею».
Довготривала пам’ять
На відміну від процесу кодування короткочасної пам’яті, що відбувається у морському конику, довготривалі спогади зберігаються у різноманітних частинах нової кори. Очевидно, різні елементи спогадів - візуальні, нюхові, слухові тощо - містяться у відповідних ділянках кори, що пов’язані з такими функціями. Ці елементи поєднуються внаслідок довготривалих змін сили передавання на відповідних синаптичних з’єднаннях таким способом, що всі компоненти образу переходять до свідомості, коли особа відтворює спогад у пам’яті.
Як тільки довготривалі спогади закладено, їх можна відтворити за допомогою значної кількості різноманітних асоціацій. Наприклад, спогад про яскраву сцену можна відновити не лише у випадку подібної сцени, а й за допомогою звуків чи запаху, пов’язаного зі сценою чи словами «сцена», «яскравий» та ін. Отже, до кожного кумульованого спогаду повинні бути численні шляхи. Крім того, багато спогадів мають емоційний компонент. Говорячи простіше, спогади можуть бути приємними чи неприємними.
Відоме та невідоме
Цікаво, що подразнення деяких ділянок скроневих часток у людей зумовлює зміни у сприйнятті навколишнього середовища. Наприклад, від дії подразника суб’єкт може почуватися незвично у відомому місці або ж відчувати: те, що стається, вже було в минулому. Виникнення такого почуття у відповідних ситуаціях, можливо, допомагає нормальній особі пристосуватися до середовища довкілля. У невідомому оточенні особа перебуває в стані тривоги й настороженості, тоді як у відомому її пильність послаблена. Неадекватне відчуття знайомості під час нових подій чи в новому оточенні клінічно відоме як явище dejа VU (у перекладі із фр. - вже побачене). Це явище трапляється від часу до часу у здорових людей, однак може виникати як аура (особливе відчуття безпосередньо перед нападом) у пацієнтів з епілепсією у випадку ураження скроневої частки.
Висновок
Отже, кодування свідомої пам’яті все ще потребує детального вивчення. Відповідно до різних поглядів інформація про відчуття тимчасово нагромаджується у різноманітних частинах передлобової ділянки кори як робоча пам’ять. Інформація в робочій пам’яті переходить до медіальної скроневої частки і, особливо, до закрутки морського коника, а звідти - до морського коника, де відбувається її опрацювання у досі нез’ясований спосіб. У цей період, як описано вище, активність є вразливою. Сигнал із морського коника переходить через підставку в напрямі нюхової кори і якимось чином зв’язує та посилює кругообіг у багатьох різних ділянках нової кори, формуючи з часом стійкі спогади, які можна відновити за допомогою багатьох різних асоціацій.
Хвороба Альцгеймера і стареча деменція
Для хвороби Альцгеймера характерною є втрата короткочасної пам’яті, що прогресує, після якої настає загальна втрата коґнітивної (від англ. congenitive - пізнавальний) функції та смерть у людей середнього віку. Подібний деґрадаційний процес в осіб старшого віку називають старечою деменцією типу Альцгеймера, однак часто також хворобою Альцгеймера. Здебільшого ця хвороба спорадична, та трапляються й сімейні типи захворювання. Вони становлять 50-60% від усіх випадків старечої деменції. Пацієнти з хворобою Альцгеймера потребують цілодобового догляду. Оскільки 10-15% населення віком понад 65 років та майже 50% населення віком понад 85 років притаманний певний рівень деменції, то такий стан є не лише серйозною медичною проблемою, а й економічною, особливо з огляду на те, що кількість старших людей серед населення розвинутих країн збільшується.
Ранніми змінами у випадку хвороби Альцгеймера є атрофія морського коника та нюхової кори. Ці зміни можна фіксувати за допомогою ЯМР упродовж двох років, перш ніж буде визначено точний діагноз. У цьому разі відбувається втрата холінергічних та інших нейронів у корі головного мозку і гостра втрата холінергічних нейронів у базальному ядрі Мейнерта й подібних ядрах, що містять тіла клітин цих нейронів, які проектують до морського коника, мигдалеподібного тіла та нової кори (див. Розділ 15). Незначних тимчасових поліпшень стану у випадку хвороби Альцгеймера інколи можна досягти за допомогою препаратів, що інгібують ацетилхолінестеразу і, відповідно, послаблюють втрату ацетилхоліну. Щоправда, таке лікування не впливає на дегенеративні процеси, що є в основі цієї хвороби.
Причина нейронної дегенерації у випадку хвороби Альцгеймера досі не з’ясована. Цитопатологічними ознаками хвороби є внутрішньоклітинні нейрофібрилярні клубочки, які складаються частково з гіперфосфорильованих форм білка тау, що в нормальному стані зв’язується з мікротрубочками (див. Розділ 1), та позаклітинних старечих бляшок, що мають серцевину з ß-амілоїда (Aß), оточеного нервовими волокнами та реактивними гліальними клітинами. Бета-амілоїди - це пентиди Aß40 та Aß42, утворені з амілоїдного прекурсорного білка (АПБ). Зокрема, Aß42 формує нерозчинні агрегати внаслідок гідролізу АПБ. У цьому разі ключовим ензимом є у-секретаза. У близько 50% ранніх випадків хвороби Альцгеймера виникають мутації генів, що відповідають за два білки, які містяться в мембранах органел клітин: пресенілін 1, кодований геном на хромосомі 14, та майже ідентичний до нього пресенілін 2, кодований геном на хромосомі 1. Обидва гіресеніліни пов’язані з активністю у-секретази. Мутації гена на хромосомі 21, кодованого АМБ, також призводять до ранніх випадків хвороби Альцгеймера. Цікаво, що пацієнти із синдромом Дауна мають додаткову хромосому 21 (трисомія 21), і відомо, що у них деменція типу Альцгеймера розвивається в ранньому віці. Якщо Aß42 акумульований позаклітинно, то він може бути токсичним для нервових клітин. Є думки, що саме це й може спричиняти хворобу Альцгеймера.
Сьогодні відомо, що естроген сповільнює прогресування хвороби Альцгеймера. Крім того, є взаємозв’язок між ізоформами аполіпопротеїну Е (АПО Е; див. Розділ 17) та віком виникнення хвороби. Ген, що відповідає за цей білок, міститься на хромосомі 19 і утворює три головні ізоформи, відмінні між собою одними чи двома амінокислотами. В осіб, гомозиготних для ізоформи АПО-4, хвороба Альцгеймера розвивається раніше, ніж у тих, хто гомозиготний для ізоформ АПО-2 та АПО-3 чи гетерозиготний для будь-якої з них. Однак для того, щоб з’ясувати роль Aß, естрогену та ізоформи АПО Е в патогенезі цієї хвороби, треба виконати значну кількість додаткових досліджень.
Цікаво, що вибіркова дегенерація в процесі старіння може відбуватися в трьох різних типах клітин у ЦНС, спричинюючи три різні хвороби, що прогресують, призводять до деформації, а в кінцевому підсумку - до смерті. Як уже зазначено, дегенерація холінергічних нейронів в основі переднього мозку пов’язана з хворобою Альцгеймера, дегенерація дофамінергічних нейронів у чорній речовині - з хворобою Паркінсона (див. Розділ 12), а дегенерація холінергічних рухових нейронів у стовбурі мозку та спинному мозку - з однією з форм бокового аміотрофічного склерозу (БАС). БАС часто називають хворобою Лу Ґеріґа, оскільки цей відомий американський бейсболіст помер від неї. Усі три хвороби переважно спорадичні, однак бувають і родинні форми. У 5% випадків БАС є родинним, у 40% із цих п’яти фіксують мутацію гена, відповідального за Сu/Zn супероксиддисмутазу (СОД-1) на хромосомі 21. Дефективний СОД-1 ген може допускати нагромадження вільних радикалів, які руйнують нейрони. Дегенеративні хвороби нервової системи, зумовлені посиленими тринуклеотидними повторюваннями, розглянуто у Розділі 12.