Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції травної системи
Травлення та всмоктування
Білки й амінокислоти
Травлення білків
Травлення білків розпочинається в шлунку, де пепсини розщеплюють деякі з пептидних зв’язків. Подібно до багатьох інших ензимів, причетних до травлення, пепсини секретуються у формі неактивних попередників (проензимів) і активуються в шлунково-кишковому тракті. Попередники пепсинів називають пепсиногенами, їх активує хлористоводнева кислота, яку виробляє шлунок. Слизова оболонка шлунка людини містить певну кількість споріднених пепсиногенів, які можна розділити на дві відмінні групи за гістохімічними ознаками: пепсиноген І та пепсиноген II. Пепсиноген І міститься тільки в кислотосекрету- вальних ділянках, тоді як пепсиноген II - також ще й у пілоричній ділянці. Максимальна секреція кислоти корелює відповідно до рівня пепсиногену І.
Таблиця 25-2. Транспортування речовин у нормі та місця максимальної абсорбції або секреції'1
Всмоктування |
Відділи тонкої кишки |
Ободова кишка |
||
проксимальні2 |
середні |
дистальні |
||
Цукри (глюкоза, галактоза та інші) |
++ |
+++ |
++ |
0 |
Амінокислоти |
++ |
+++ |
++ |
0 |
Водо- та жиророзчинні вітаміни, за винятком В12 |
+++ |
++ |
0 |
0 |
Бетаїн, диметилгліцин, саркозин |
+ |
++ |
++ |
? |
Антитіла у новонароджених |
+ |
++ |
+++ |
? |
Піримідини (тимін та урацил) |
+ |
+ |
? |
? |
Абсорбція жирних кислот з довгими ланцюгами та |
+++ |
++ |
+ |
0 |
перетворення у тригліцериди |
||||
Жовчні солі |
+ |
+ |
+++ |
|
Вітамін В12 |
0 |
+ |
+++ |
0 |
Na+ |
+++ |
++ |
+++ |
+++ |
К+ |
+ |
+ |
+ |
Сек |
Са2+ |
+++ |
++ |
+ |
? |
Fe2+ |
+++ |
++ |
+ |
? |
Cl- |
+++ |
++ |
+ |
+ |
SO42- |
++ |
+ |
0 |
? |
1 Рівень абсорбції позначено від + до +++. Сек - секретується, якщо К+ у просвіті кишки є низьким.
2 До проксимальних частин тонкої кишки належить голодна та дванадцятипала кишка (крім зауваженого винятку, коли дванадцятипала кишка секретує НСO3 - і має незначну кінцеву абсорбцію або секрецію NaCl).
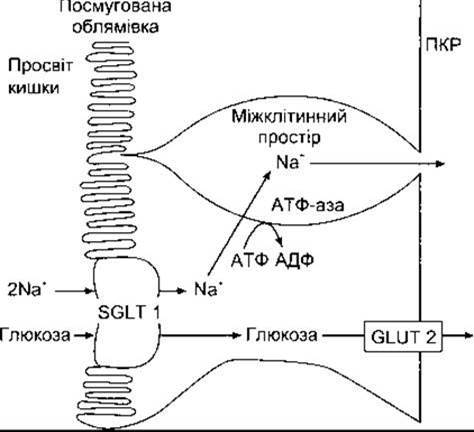
Рис. 25-3. Механізм транспортування глюкози через кишковий епітелій. Транспортування глюкози до ентероцитів поєднане з транспортуванням Na+ і використанням котранспортера SGLT 1. Потім Na+ активно переходить з клітини, і глюкоза потрапляє в інтерстиційний простір шляхом полегшеної дифузії через GLUT 2, а звідси згодом дифундує у кров.
Пепсини гідролізують зв’язки між ароматичними амінокислотами, такими як фенілаланін або тирозин, та іншою амінокислотою, отже, продуктами пептичного травлення є поліпептиди різноманітних розмірів. У шлунку також міститься желатиназа, яка перетворює в рідкий стан желатин. Хімозин - шлунковий ензим, що спричинює зсідання молока, відомий також як ренін - міститься в шлунку молодих тварин; інколи його немає у людей.
Оскільки пепсини оптимально активні при pH 1,6-3,2, то їхня дія завершується, коли шлунковий вміст змішується з лужним панкреатичним соком у дванадцятипалій та голодній кишці. Значення pH кишкового вмісту в цибулині дванадцятипалої кишки 2,0-4,0, проте в інших частинах - близько 6,5.
У тонкій кишці поліпептиди, що утворились під час протеолізу в шлунку, перетравлюються за допомогою високоактивних протеолітичних ензимів підшлункової залози та слизової оболонки кишки. Трипсин, хемотрипсин, еластаза гідролізують внутрішні пептидні зв’язки в пептидних молекулах, тому їх називають ендопептидазами. Утворення активних ендопептидаз з їхніх неактивних попередників описано у Розділі 26. Карбоксипептидази підшлункової залози - це екзопептидази, які гідролізують амінокислоти в карбоксильних та амінокінцевих групах поліпептидів. Деякі вільні амінокислоти вивільняються у кишковий просвіт, тоді як інші - у вільному стані на поверхню клітин після попереднього розщеплення амінопептидазами, карбоксипептидазами, ендопептидазами та дипептидазами щіточкової облямівки слизових клітин. Деякі ди- та трипептиди активно переходять до кишкових клітин, їхні внутрішньоклітинні гіептидази гідролізують до амінокислот, які згодом потрапляють до кров’яного русла. Отже, остаточне травлення білків до амінокислот відбувається в трьох місцях: кишковому просвіті, щіточковій облямівці та цитоплазмі слизових клітин.
Всмоктування
В ентероцитах є сім відмінних транспортних систем для переміщення амінокислот. П’ять з них потребують наявності йонів Na+, крім того, котранспорт амінокислот і Na+ відбувається подібно до котранспорту Na+ і глюкози (див. рис. 25-3). Дві транспортні системи також потребують СІ. У двох системах транспортування залежить від Na+.
Ди- та трипептиди переходять до ентероцитів системою, що потребує Н+ замість на Na+. Це так зване мале всмоктування великих пептидів. В ентероцитах амінокислоти вивільняються з пептидів шляхом внутрішньоклітинного гідролізу, крім того, амінокислоти, що всмоктуються з кишкового просвіту та щіточкової облямівки, транспортуються назовні від ентероцитів через їхній базолатеральний край за допомогою п’яти транспортних систем, а звідси потрапляють у кров’яне русло печінкової ворітної вени. Дві з цих транспортних систем залежать від Na+, а решта - ні. Значна кількість малих пептидів також належать до портальної системи.
Всмоктування амінокислот у дванадцятипалій та голодній кишці відбувається швидко, а у клубовій - повільніше. Зі спожитою їжею надходить близько 50% від усіх перетравлених білків, з білків травних соків - 25%, зі злущених клітин слизової - 25%. Тільки 2-5% білків у тонкій кишці не піддаються травленню та всмоктуванню. Деяка частина від поглинутих білків потрапляє до ободової кишки і зрештою їх перетравлюють бактерії. Наявність білка у фекальних випорожненнях є не харчового походження, а наслідком руйнування бактерій та клітин. Про це свідчать випадки, коли пептидазна активність щіточкової облямівки і цитоплазми клітин слизової збільшується після резекції частини клубової кишки або незалежного ураження під час голодування. Отже, ці ензими є суб’єктами гомеостатичного регулювання. У людей вроджений дефект механізму, що транспортує нейтральні амінокислоти в кишці та ниркових канальцях, спричинює хворобу Хартнуна. Вроджений дефект транспортування головних амінокислот зумовлює цистинурію.
У новонароджених помірна кількість неперетравлених білків також може всмоктуватися. Білкові антитіла в материнському молозиві - це великі секреторні імуноглобуліни (IgA), які за допомогою трансцитозу проходять через епітелій молочної залози; їхнє утворення збільшується в молочній залозі наприкінці вагітності. У разі грудного вигодовування IgA всмоктуються з кишки в систему кровообігу немовлят і забезпечують пасивний імунітет проти інфекцій. Всмоктування відбувається шляхом ендоцитозу з наступним екзоцитозом.
З віком всмоктування білків послаблюється, хоча й у дорослих простежується в незначних кількостях. Сторонні білки, що потрапляють у кровоплин, зумовлюють утворення антитіл і реакції антиген-антитіло, які трапляються у випадку повторного потрапляння більшої кількості того ж самого білка, спричинюють алергічні симптоми після споживання певного виду їжі. Поширеність харчової алергії у дітей перевищує 8%. Деяка їжа є сильнішим алерген ним чинником порівняно з іншою. Ракоподібні, молюски, риба - загальновідомі алергени, хоча досить часто простежуються алергічні відповіді на коров’яче молоко, яйця та різні делікатеси.
Всмоктування білкових антигенів, особливо бактеріальних та вірусних білків відбувається у великих М-клітинах (від англ. microfold - мікроскладка) спеціалізованих кишкових епітеліоцитах, що містяться у скупчених лімфатичних вузликах (плямках Пеєра). Ці клітини передають антигени до лімфоїдних тканин, після чого розпочинається активування лімфобластів. Активовані лімфобласти потрапляють у систему кровообігу, а пізніше повертаються до слизової оболонки кишки й інших епітеліоцитів, де секретують IgA у відповідь на повторну появу того ж самого антигену. Такий секреторний імунітет є важливим захисним механізмом, він детально описаний у Розділі 27.
Нуклеїнові кислоти
У кишці нуклеїнові кислоти за допомогою панкреатичних нуклеаз розщеплюються на нуклеотиди, а ті в подальшому за участю ензимів, що містяться на люмінальній мембрані слизових клітин, - на нуклеозиди та фосфорну кислоту. Нуклеозиди, відповідно, розщеплюються на такі складові: цукри і пуринові та пірамідинові основи, що їх абсорбує активне транспортування.