Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції травної системи
Механізми регулювання шлунково-кишкових функцій
Екзокринна частина підшлункової залози
Підшлунковий сік уміщує ензими, які мають важливе значення для травлення (див. табл. 25-1). Його секрецію регулює частково рефлекторний механізм і частково гастроінтестинальні гормони секретину та ХЦК.
Анатомічні особливості
Частина підшлункової залози, що секретує підшлунковий сік, складається з альвеолярної тканини, що подібна до слинних залоз. Гранули вміщують травні ензими (зимогенні гранули), які утворюються у клітинах і які вивільняє екзоцитоз (див. Розділ 1) з верхівок клітин у просвіт панкреатичних проток (рис. 26-16). Міжчасточкові вивідні протоки об’єднуються у загальну вивідну протоку (протока Вірсунґа), яка звичайно з’єднується із загальною жовчною протокою, утворюючи печінково-панкреатичну ампулу Фатера (рис. 26-17). Ампула відкривається через дуоденальну пипку, і її устя оточене сфінктером Одді. У деяких осіб є додаткова панкреатична протока (протока Санторіні), яка долучується до дванадцятипалої кишки проксимальніше.
Склад підшлункового соку
Підшлунковий сік має лужну реакцію (табл. 26-6), оскільки вміщує високу концентрацію НСО3 (приблизно 113 мекв/л, у плазмі - 24 мекв/л). Понад 1500 мл підшлункового соку секретовано щодня. Жовч та кишковий сік також є нейтральними або лужними і разом ці три секрети нейтралізують шлункову кислотність, збільшуючи pH дуоденального вмісту до 6,0-7,0. На момент надходження хімусу у голодну кишку кислотно-лужна реакція стає майже нейтральною, проте інтестинальний вміст зрідка буває лужним.
Таблиця 26-6. Склад підшлункового соку у нормі
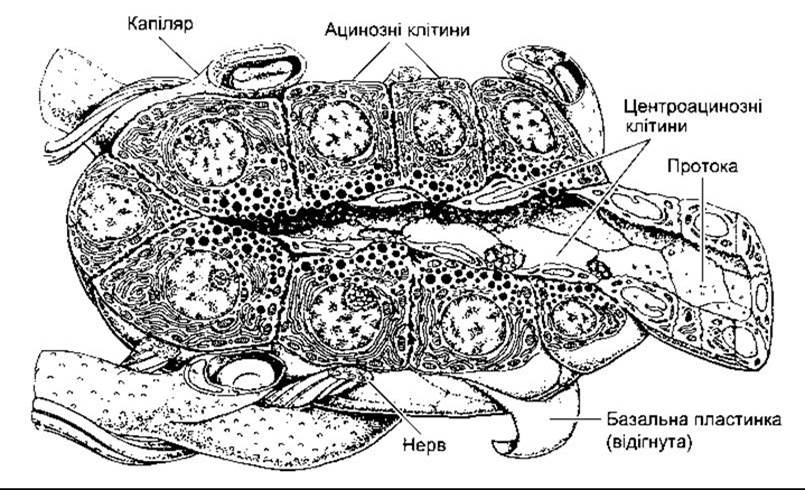
Рис. 26-16. Ацинозні клітини, що оточують кінцеву частину панреатичних проток. Зверніть увагу на багату ґранулярну (шорстку) ендоплазматичну сітку та ґранули зимогену, що концентровані на верхівках клітин (відтворено за дозволом з Krstic RV: Die Gewebe des Menschen und der Saugetiere. Springer, 1978).
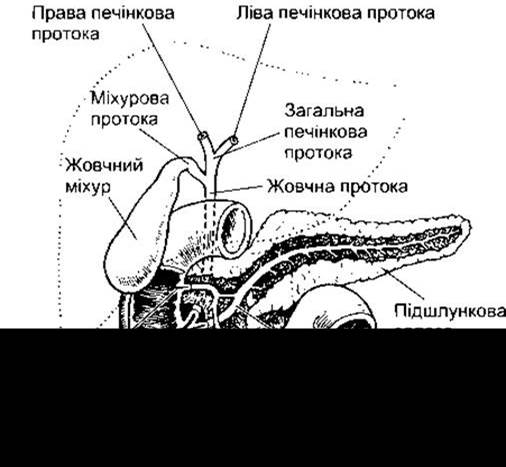
Рис. 26-17. З’єднання проток жовчного міхура, печінки та підшлункової залози (відтворено за дозволом з Bell GH, Emslie-Smith D, Paterson CR. Textbook of Physiology and Biochemistry, 9th ed. Churchill Livingstone, 1976).
Потужні білковорозщеплювальні ензими панкреатичного соку секретовані як неактивні проензими. Трипсиноген перетворюється в активний ензим трипсин за допомогою ензиму посмугованої облямівки ентеропептидази (ентерокінази) в разі надходження панкреатичного соку у дванадцятипалу кишку. Ентеропептидаза вміщує 41% полісахаридів, і такий високий вміст полісахаридів попереджує його самоперетравлення перш ніж він може вплинути. Трипсин перетворює хімотрипсиногени у хімотрипсини, а інші проензими в активні ензими (рис. 26-18). Він також може активувати трипсиноген; однак утворюється тільки один трипсин і це є автокаталітична ланцюгова реакція. Дефіцит ентеропептидази трапляється як вроджена аномалія і призводить до білкового порушення харчування.
Безперечно, потенційно небезпечним є вивільнення у підшлункову залозу малих порцій трипсину. Внаслідок ланцюгових реакцій можуть активізуватися активні ензими, здатні до самоперетравлення підшлункової залози, тому не випадково підшлункова залоза у нормі вміщує інгібітор трипсину.
Іншим ензимом, що активує трипсин, є фосфоліпаза А2. Цей ензим розщеплює жирні кислоти від лецитину, утворюючи лізолецитин, який ушкоджує клітинні мембрани. Є гіпотеза, що в разі гострого панкреатиту, важкого, а інколи фатального захворювання, фосфоліпаза А2 активує панкреатичні протоки до утворення лізолецитину з лецітину, який у нормі є складовим компонентом жовчі. Це спричинює розрив тканини підшлункової залози і некроз навколишньої жирової тканини.
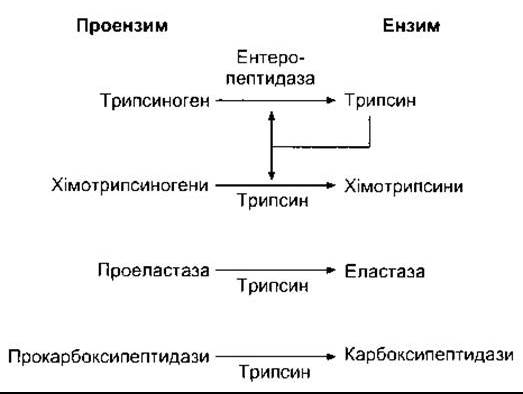
Рис. 26-18. Активування панкреатичних протеаз у дванадцятипалій кишці
Невеликі кількості панкреатичних травних ензимів потрапляють у кровообіг, однак у разі гострого панкреатиту рівень травних ензимів, що циркулюють у крові, значно підвищується. Вимірювання концентрації амілази або ліпази плазми є діагностичним критерієм захворювання.
Регулювання секреції підшлункового соку
Секрецію підшлункового соку початково регулюють гуморальні механізми. Секретин впливає на панкреатичні протоки, спричинюючи значну секрецію лужного підшлункового соку, багатого на НСО3 і бідного на ензими. Вплив на клітини проток виявляється у збільшенні внутрішньоклітинної цАМФ. Секретин також стимулює секрецію жовчі. ХЦК діє на ацинарні клітини, спричинюючи вивільнення гранул зимогену й утворення підшлункового соку, багатого на ензими, однак невеликого об’єму. Цей вплив стимулює фосфоліпаза С (див. Розділ І).
Відповідь на внутрішньовенне введення секретину зображена на рис. 26-19. Зверніть увагу, що об’єм підшлункового соку збільшується, концентрація Сl у ньому зменшується, а концентрація НСO3 збільшується. Хоча НСО3 секретують малі протоки, проте реабсорбування відбувається у великих протоках в обмін на Сl. Інтенсивність обміну протилежно пропорційна до швидкості, з якою витікає підшлунковий сік.
Подібно до ХЦК ацетилхолін впливає на ацинарні клітини через фосфоліпазу С, сприяючи вивільненню гранул зимогену, а стимулювання блукаючого нерва спричинює секрецію невеликої кількості підшлункового соку, багатого на ензими. Це є свідченням умовного рефлексу, забезпечуваного блукаючим нервом у відповідь на вигляд або запах їжі.
Епітелій малих вивідних проток підшлункової залози містить багато CFTRs (див. Розділ І). Зв’язок CFTRs з секрецією НСО3 остаточно невідомий, однак у разі цистичного фіброзу (муковісцидозу) вміст НСО3 та об’єм панкреатичного соку зменшуються, а кількість випадків хронічних панкреатитів збільшується.
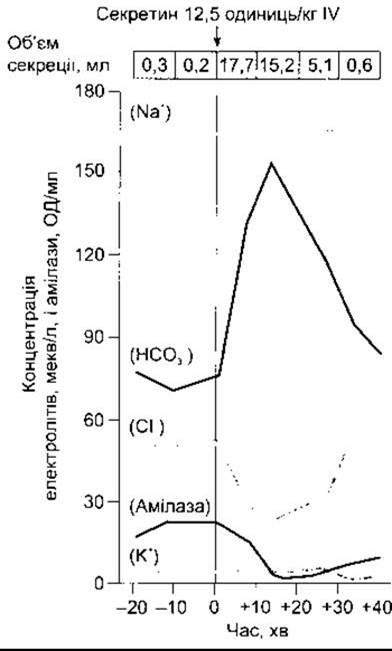
Рис. 26-19. Вплив одноразової дози секретину на склад та об’єм підшлункового соку в людей (модифіковано і відтворено за дозволом з Janowitz HD: Pancreatic secretyon. Physiol Physicians [Nov] 1964;2:11).