Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Збудлива тканина: м'язова
Гладкі м'язи - Вісцеральні гладкі м'язи
Електрична та механічна активність
Вісцеральні гладкі м’язи мають нестабільний мембранний потенціал, їм властиві хаотичні скорочення, які не залежать від впливу іннервації. Такий стан постійного неповного скорочення називають тонусом. Для мембранного потенціалу вісцеральних гладких м’язів не характерне таке поняття, як потенціал спокою, оскільки мембранний потенціал порівняно низький в активному стані тканини, і високий у разі гальмування, проте під час періодів спокою він становить близько -50 мВ. На мембранний потенціал накладаються різноманітні електричні хвилі (рис. 3-17). Ці хвилі є повільними синусоїдними коливаннями з амплітудою декілька мілівольтів і піками, які інколи переходять через нульову відмітку потенціалу дії, а деколи - ні. У більшості тканин піки тривають близько 50 мс. Однак для деяких тканин характерні подовжені плато під час фази реполяризації, подібно до потенціалу дії міокарда. Піки можуть простежуватися як під час фази підіймання, так і під час фази спадання синусоїдних коливань. Крім того, у гладких м’язах утворюються пейсмейкерні потенціали, подібні до тих, що їх генерують аналогічні серцеві пейсмейкерні клітини. Проте у вісцеральних гладких м’язах ці потенціали утворюються у багатьох ділянках, розташування яких постійно змінюється. Піки, що генеруються в пейсмейкерних клітинах, поширюються у м’язі на певну відстань. Оскільки м’яз постійно перебуває в активному стані, то вивчати зв’язки між електричними та механічними явищами у вісцеральних гладких м’язах досить важко. Поодинокі електричні піки можна одержати у деяких препаратах порівняно малоактивних м’язів. М’яз скорочується через близько 200 мс від початку піка і за 150 мс після його закінчення. Максимальне напруження в разі скорочення буде лише через 500 мс від початку піка. Отже, координація збудження та скорочення у вісцеральних гладких м’язах є дуже повільним процесом, на відміну від такого процесу у скелетних м’язах і міокарді, де інтервал між початком деполяризації та ініціацією скорочення становить до 10 мс.
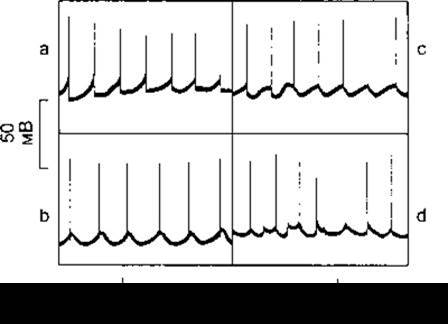
Рис. 3-17. Спонтанна електрична активність окремих гладком’язових клітин товстої кишки мурчака: а - пейсмейкерний тип; b - синусоїдні хвилі з потенціалами дії у фазі підіймання; с - синусоїдні хвилі з потенціалами дії у фазі спадання; d - суміш пейсмейкерних, синусоїдних електричних коливань та потенціалів дії (відтворено за дозволом з Bulbring Е: Physiology and pharmacology of intestinal colon smooth muscle. Lectures on the Scientific Basis of Medicine 1957;7:374).
Молекулярні основи м’язового скорочення
Як і в скелетних м’язах, ініціація скорочення гладких м’язів відбувається завдяки Са2+. Проте вісцеральні гладкі м’язи мають погано розвинену саркоплазматичну сітку. Тому збільшення концентрації Са2+ всередині клітини, яке є безпосередньою причиною скорочення, виникає головно внаслідок надходження Са2+ з ПКР через потенціалозалежні Ca2+-канали. Крім того, у гладких м’язах для активування міозинової АТФ-ази необхідне фосфорилювання самої молекули міозину. Фосфорилювання та дефосфорилювання міозину відбувається також і в скелетних м’язах, однак воно не є обов’язковою передумовою активування АТФ-ази. У гладких м’язах Са2+ зв’язується з кальмодуліном (див. Розділ 1), і комплекс, що утворюється внаслідок цього, активує кальмодулінозалежну кіназу легких ланцюгів міозину. Цей ензим є каталізатором реакції фосфорилювання легких ланцюгів міозину, під час якої фосфатний залишок приєднується до серину в положенні 19. Фосфорилювання забезпечує подальше активування АТФ-ази, і молекули актину починають ковзати по міозинових ланцюгах, спричинюючи скорочення м’яза. Цей процес цілком протилежний до того, що відбувається у скелетних м’язах та міокарді, де скорочення спричинене зв’язуванням Са2+ з тропоніном С.
Дефосфорилювання міозину зумовлює міозин-фосфатаза. Цей ензим у фосфорильованому стані інгібований, а його активування відбувається внаслідок дефосфорилювання. Його дефосфорилює rho-асоційована кіназа, яку, відповідно, активують ліганди, що пригнічують активність гладкої мускулатури. Проте, дефосфорилювання кінази легких ланцюгів міозину не обов’язково призводить до розслаблення гладких м’язів. Для гладких м’язів характерне утворення так званих поперечних містків, тобто міозин зв’язаний з актином упродовж деякого часу після зменшення концентрації Са2+ у цитоплазмі. Це явище забезпечує тривале скорочення із незначними витратами енергії, що особливо важливо для гладкої мускулатури судин. Уважають, що м’яз розслаблюється тоді, коли відбулась остаточна дисоціація Са2+-кальмодулінового комплексу або коли почали діяти інші механізми. Головні процеси, що відбуваються під час скорочення і розслаблення вісцеральних гладких м’язів, наведені в табл. 3-6. У поліелементних гладких м’язах відбуваються загалом ідентичні процеси.
Варто описати відмінності між серцевим та гладкими м’язами, оскільки обидва ці типи м’язової тканини утворюють серцево-судинну систему. Скоротливі відповіді міокарда мають фазовий характер із чітким чергуванням скорочень та розслаблень, тоді як скорочення гладких м’язів часто є тонічними завдяки утворенню поперечних містків. Підвищення внутрішньоклітинної концентрації цАМФ збільшує силу скорочень серцевого м’яза, водночас на гладкі м’язи цАМФ впливає зворотним чином - розслабляє їх, оскільки інгібує реакцію фосфорилювання кінази легких ланцюгів міозину.
Подразнення
Вісцеральні гладкі м’язи мають одну унікальну властивість, якої нема в усіх інших типів м’язової тканини. Вони скорочуються в разі розтягування без будь-якої іннервації. Реагуванням на розтяг є зменшення мембранного потенціалу, збільшення частоти піків і посилення загального тонусу м’язів.
Якщо in vitro до середовища, в якому перебуває препарат гладкої мускулатури стінки кишки, додати адреналін або норадреналін, то мембранний потенціал збільшиться, частота піків стане меншою, і м’яз розслабиться (рис. 3- 18). Норадреналін - це хімічний трансмітер, що його вивільнюють норадренергічні нервові закінчення (див. Розділ 4); подразнення норадренергічних нервів чинить гальмівний вплив на гладкі м’язи (див. Розділ 4). Стимуляція in vivo норадренергічних нервів, які проходять до кишки, зумовлює послаблення її активності. Норадреналін впливає як на а-, так і на ß-адренергічні рецептори м’язів (див. Розділ 4). Вплив на ß-рецептори виявляється зниженням тонусу м’язів у відповідь на подразнення. Трансмітером цього ефекту є цАМФ (див. Розділ 1), а безпосередньою причиною - збільшення внутрішньоклітинного зв’язування Са2+. Вплив на а-адренорецептори, що також виявляється гальмуванням скорочення, зумовлений посиленням виходу йонів Са2+ за межі м’язових клітин.
Вплив ацетилхоліну на мембранний потенціал та скоротливу активність гладких м’язів цілком протилежний до дії норадреналіну. Якщо ацетилхолін додати в рідину, у якій міститься препарат гладкого м’яза in vitro, то мембранний потенціал зменшиться, і піки стануть частішими (див. рис. 3-18). М’яз стане активнішим, посилиться тонічне напруження і збільшиться кількість ритмічних скорочень. Медіаторами цього ефекту є фосфоліпаза С та ІР3 які збільшують концентрацію Са2+ всередині клітини. У інтактних тварин стимулювання холінергічних нервів спричинює вивільнення ацетилхоліну, виникнення збудливих потенціалів (див. Розділ 4) та посилення перистальтики кишки. Подібні ж ефекти in vitro зумовлені дією холоду та розтягу.
Функція нервів гладкої мускулатури
Таблиця 3-6. Послідовність явищ у разі скорочення і розслаблення вісцеральних гладких м’язів1

1 Порівняйте з табл. 3-2.
Вплив ацетилхоліну та норадреналіну відображає дві важливі риси гладких м’язів: по-перше, їхню спонтанну активність без будь-якого нервового подразнення; по-друге, чутливість до різноманітних хімічних чинників, що вивільняються з нервових закінчень або містяться у кровоплині. У ссавців вісцеральні м’язи, як звичайно, мають подвійну іннервацію з двох відділів автономної нервової системи. Структура та функції сполучень між цими нервами та гладкими м’язами описані у Розділі 4. Завданням нервово-регуляторних механізмів є не ініціювання активності гладких м’язів, а радше, модулювальний вплив на нього. Стимулювання одного відділу автономної нервової системи посилює активність гладких м’язів, тоді як стимулювання іншого - послаблює її. У деяких органах норадренергічне стимулювання посилює, а холінергічне - послаблює активність гладкої мускулатури, для інших же характерні протилежні властивості.
Зв’язок між довжиною та напруженням. Пластичність
Іншою специфічною властивістю гладких м’язів є варіабельність напруження, що виникає у разі заданої довжини. Якщо шматочок вісцерального гладкого м’яза розтягнути, то він спочатку виявляє збільшення напруження (див. вище). Однак якщо м’яз утримувати в розтягнутому положенні, то напруження у ньому поступово спадає. Деколи воно стає таким же або навіть меншим від того, що спостерігали до розтягнення. Практично неможливо з’ясувати зв’язок між довжиною і напруженням гладкого м’яза. Для нього нема такого поняття, як довжина спокою. Певною мірою гладкі м’язи більше нагадують в’язку масу, аніж тканину пружної структури. Цю властивість називають пластичністю гладкої мускулатури.
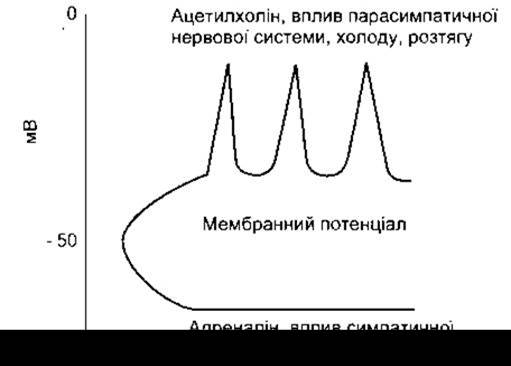
Рис. 3-18. Вплив різноманітних чинників на мембранний потенціал гладких м’язів кишки.
Фізіологічне значення пластичності можна відобразити у дослідах на здорових людях. Наприклад, можна вимірювати напруження, що виникає в гладком’язових стінках сечового міхура за різних ступенів розтягу (рідину в цьому випадку вводять у сечовий міхур через катетер; див. рис. 38-27). На початку в разі збільшення об’єму простежується порівняно незначне зростання напруження завдяки пластичності стінок сечового міхура, однак згодом у деякий момент сечовий міхур із силою скорочується.