Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Синаптичне і контактне передавання
Синаптичне передавання - Електричні явища в постсинаптичних нейронах
Методика, що дає змогу проникнути в клітину спинного мозку, допомагає вивчати явища електричної активності в постсинаптичному нейроні. Цього досягають шляхом уведення мікроелектрода в вентральну частину спинного мозку. Прокол клітинної мембрани ідентифікують за виникненням стійкої (-70 мВ) різниці потенціалів між мікро- електродом і електродом, що розміщений поза клітиною. Підтвердженням того, що клітина є спинномозковим мотонейроном, слугує подразнення відповідного переднього корінця і наступне фіксування електричної активності клітини. Таке подразнення зумовлене антидромним імпульсом (див. Розділ 2), який поширюється до тіла клітини і зупиняється в цій точці. Отже, наявність потенціалу дії в клітині після антидромного подразнення свідчить про те, що клітина, у яку введено мікроелектрод, справді є мотонейроном спинного мозку, а не інтернейроном. Активність у певних пресинаптичних закінченнях, що контактують з клітиною, у яку введено мікроелектрод, може бути ініційована шляхом подразнення заднього корінця (рис. 4-7).
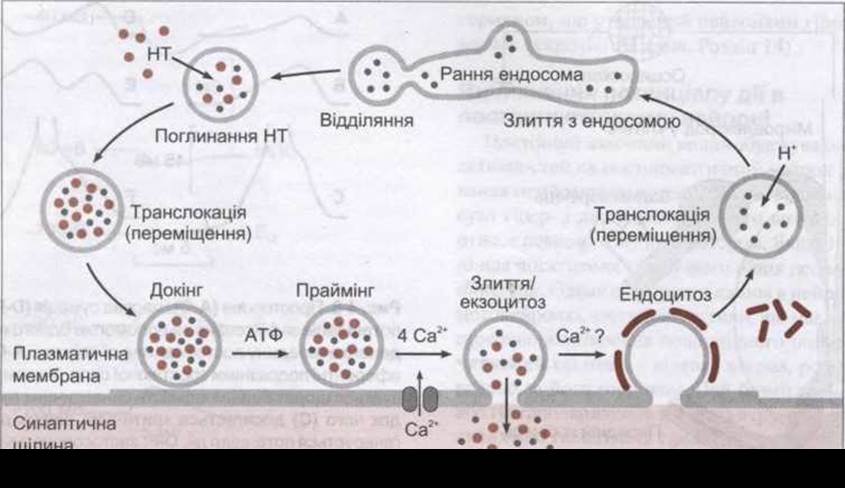
Рис. 4-5. Цикл маленьких синаптичних пухирців у пресинаптичному нервовому закінченні. Пухирці відділяються від ранньої ендосоми і наповнюються нейротрансмітером (НТ; угорі ліворуч). Далі вони рухаються до плазматичної мембрани, після чого і відбувається їхній докінг і праймінг. У випадку надходження до закінчення потенціалу дії, надходження Са2+ зумовлює процеси злиття й екзоцитозу, і вміст пухирця виділяється в синаптичну щілину. У подальшому стінка пухирця вкривається клатрином і захоплюється внаслідок ендоцитозу. В цитоплазмі вона зливається з ранньою ендосомою і цикл повторюється (відтворено за дозволом з Sudhof ТС: The synaptic vesicle cycle: a cascade of protein-protein interactions. Nature 1995:375:645).
Збуджувальні постсинаптичні потенціали
Поодинокі подразнення чутливих нервів за умов експериментів, описаних вище, не призводять до появи в постсинаптичному нейроні потенціалу дії, здатного поширюватись. Замість цього подразнення зумовлює або короткочасну часткову деполяризацію, або короткочасну гіперполяризацію.
Реакція деполяризації, спричинена поодиноким відповідної сили подразником, розпочинається через 0,5 мс після надходження імпульсу в спинний мозок, досягає максимуму через 1,0-1,5 мс і згодом експоненціально припиняється. Протягом цього потенціалу збудливість нейрона щодо іншого подразника зростає, внаслідок чого цей потенціал названо збуджувальним постсинаптичним потенціалом (ЗПСП). ЗПСП утворюється шляхом деполяризації постсинаптичної клітинної мембрани, суміжної з пресинаптичним закінченням. Збуджувальний трансмітер відкриває Na+- або Са2+-канали постсинаптичної мембрани, чим зумовлює виникнення внутрішнього струму. Ділянка утвореного таким способом струму є настільки маленькою, що він не поширюється, доки позитивні заряди не деполяризують цілу мембрану. ЗПСП, зумовлені активністю в одному синаптичному пухирці, невеликі, проте деполяризації, що виникають у кожному з активних синаптичних пухирців, додаються.
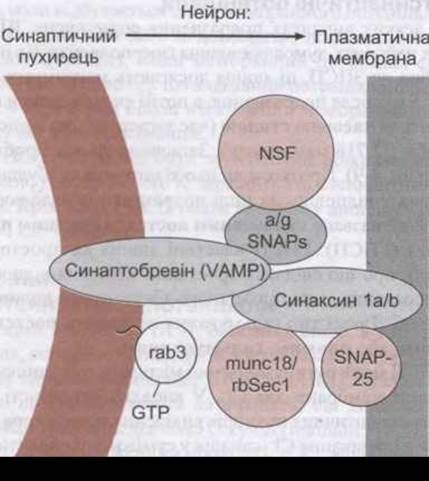
Рис. 4-6. Головні білки, що взаємодіють, зумовлюючи акцептування і злиття синаптичних пухирців нервових закінчень (відтворено за дозволом з Ferro-Novick S, John R: Vesicle fusion from yeast to man. Nature 1994:370:191).
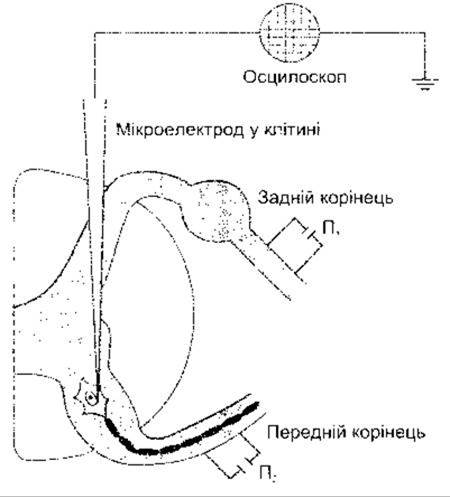
Рис. 4-7. Розміщення реєструвального і подразнювального електродів під час дослідження синаптичної активності в спинномозкових мотонейронах ссавців. Подразнення П2 використовують для виникнення антидромних імпульсів, що дає змогу ідентифікувати клітину, а подразнення П1 - для ортодромного подразнення через рефлекторну дугу.
Сумація може бути просторовою або часовою. Якщо активність виникає одночасно в більше ніж одному синаптичному пухирці, то простежується просторова сумація, і активність одного синаптичного пухирця підсилює активність іншого, наближаючись до критичного рівня деполяризації. Часова сумація виникає, коли повторні аферентні подразники зумовлюють нові ЗПСП, перш ніж згасають попередні. Очевидно, що чим довше триває ЗПСП, тим більша можливість виникнення сумації. Явища просторового і часового підсилення відображені на рис. 4-8. Отже, ЗПСП не підлягає законові «усе або нічого», а його значення пропорційне до сили аферентних подразників. Якщо ЗПСП у разі досягнення критичного рівня деполяризації в клітині є значним, то виникає повноцінний потенціал дії.
Синаптична затримка
Між часом надходження імпульсу до пресинаптичного закінчення і початком реакції в постсинаптичному нейроні виникає перерва тривалістю 0,5 мс, яку називають синаптичною затримкою. Затримка в умовах максимального подразнення пресинаптичного нейрона відповідає латентності ЗПСП і зумовлена часом, що потрібний для виділення синаптичного трансмітера і його дії на мембрану постсинаптичної клітини. Внаслідок цього час, затрачений на передавання імпульсу в ланцюгу нейронів, що містить велику кількість синапсів, є тривалішим, ніж у ланцюгу, де синапсів лише декілька. Оскільки відомо, що мінімальний час, потрібний для передавання через один синапс, становить 0,5 мс, то шляхом вимірювання часу затримки в передаванні від заднього корінця через спинний мозок до переднього корінця можна з’ясувати: є задана рефлекторна дуга моносинаптичною чи полісинаптичною (тобто містить більше одного синапсу).
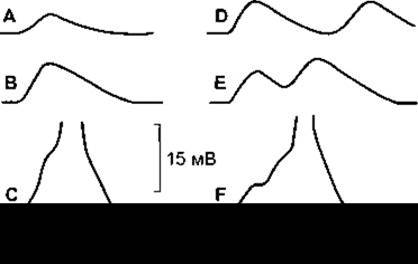
Рис. 4-8. Просторова (А-С) і часова сумація (D-F) ЗПСП. Зміни потенціалів зафіксовано за допомогою одного електрода, уведеного всередину постсинаптичної клітини. А-С: застосовано аферентні подразники зростаючої сили. У цьому випадку активується щораз більша кількість синаптичних пухирців, унаслідок чого (С) досягається критичний рівень деполяризації і ґенерується потенціал дії. D-F: застосовано два різні, однакові за силою, подразники. У випадку F часовий інтервал між ними зменшено, внаслідок чого теж досягається критичний рівень деполяризації і ґенерується потенціал дії.
Гальмівні постсинаптичні потенціали
У деяких випадках подразнення спричинене ЗПСП в одних нейронах, зумовлює явища гіперполяризації в інших. Подібно до ЗПСП, ці явища досягають максимуму через 1,0-1,5 мс після подразнення, а потім експоненціально загасають за часовою сталою (час загасання, що відповідає 1/е; або 1/2,718 максимуму). Загасання триває приблизно З мс (рис. 4-9). Протягом дії цього потенціалу збудливість нейрона у відповідь на інші подразники послаблюється, тому його названо гальмівним постсинаптичним потенціалом (ГПСП). ГПСП властиві явища як просторової сумації, про що свідчить зростання значення відповіді у разі зростання сили гальмівного аферентного впливу, так і часової. Такий тип гальмування називають постсинаптичним, або прямим, гальмуванням.
ГПСП може бути зумовлений місцевим зростанням проникності мембрани для Сl-. У випадку активності гальмівних синаптичних пухирців виділення трансмітера спричинює відкривання Сl- -каналів у суміжній зі синаптичним пухирцем ділянці постсинаптичної мембрани; Сl- переходить за концентраційним градієнтом, унаслідок чого негативний заряд переміщується всередину клітини, і мембранний потенціал збільшується. Послаблення збудливості нервової клітини під час ГПСП зумовлене зміщенням мембранного потенціалу в протилежному щодо критичного рівня деполяризації напрямі. Внаслідок цього для досягнення критичного рівня деполяризації необхідна набагато більша збуджувальна (деполяризувальна) активність. Факт, що ГПСП зумовлений Сl-, можна довести в досліді з повторним подразненням, коли відбувається зміна мембранного потенціалу спокою й утримання його за допомогою потенціалозалежної фіксації (кліпси). Коли мембранний потенціал спокою перебуває на рівні ЕС1, то гальмівний потенціал зникає (див. рис. 4-9), а в разі більш негативного мембранного потенціалу він стає позитивним.
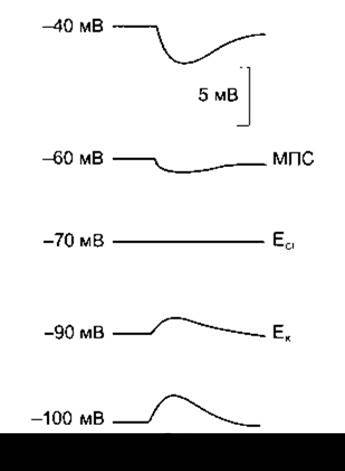
Рис. 4-9. ГПСП, зумовлений збільшенням надходження Cl-, що виникає внаслідок подразнення в умовах, коли мембранний потенціал досягає різних рівнів за допомогою потенціалозалежної фіксації; МПС - мембранний потенціал спокою нейрона. Зверніть увагу, що у випадку досягнення мембранним потенціалом рівня, який відповідає ЕСІ, ГПСП зникає, а в умовах найвищого мембранного потенціалу стає позитивним.
ГПСП теж може бути зумовлений відкриванням К+ - каналів, унаслідок чого К+ виходить з постсинаптичної клітини. Крім того, ГПСП також виникає у випадку закриття Na+ - і Са2+ -каналів.
Повільні постсинаптичні потенціали
Окрім ЗПСП та ГПСП, описаних вище, в автономних вузлах, серцевому і гладких м’язах, а також у кіркових нейронах виникають повільні ЗПСП та ГПСП. Ці постсинаптичні потенціали мають латентність, що дорівнює 100—500 мс, і тривають декілька секунд. Повільні ЗПСП зумовлені головно зниженням провідності К+, а повільні ГПСП - її підвищенням.
У симпатичних вузлах виявлено також пізні повільні збуджувальні постсинаптичні потенціали з латентністю в 1-5 с і тривалістю 10-30 хв. Цей потенціал теж зумовлений, принаймні частково, зниженням провідності К+, а трансмітером цього є пептид, тісно пов’язаний з ГнРГ - гормоном, що утворений нейронами гіпоталамуса і стимулює секрецію ЛГ (див. Розділ 14).
Виникнення потенціалу дії в постсинаптичному нейроні
Постійний взаємний вплив збуджувальної і гальмівної активностей на постсинаптичний нейрон зумовлює коливання мембранного потенціалу, що відповідає алгебричній сумі гіпер- і деполяризувального впливів. Тіло нейрона, отже, є певного типу інтегратором. Якщо 10-15 мВ достатні для досягнення критичного рівня деполяризації, то виникає пік. Однак електричні явища в нейроні складніші. В мотонейронах частиною клітини, що має найнижчий поріг стосовно виникнення повноцінного потенціалу дії, є початковий сегмент - ділянка аксона, розміщена безпосередньо за його горбиком. Цей безмієліновий сегмент де- або гіперполяризують джерела струму, що виникає в збудливих або гальмівних синаптичних пухирцях. Ця ділянка нейрона активізується першою, а далі потенціал поширюється в двох напрямах - уздовж аксона і в зворотному напрямі, тобто до тіла клітини. Ретроградні імпульси, що досягають тіла клітини, ймовірно, важливі для «очищення» від попередніх впливів, що необхідно для наступного повторення взаємного впливу на клітину збуджувальної та гальмівної активностей.
Функція дендритів
Тривалий час уважали, що дендрити - це ділянки, де виникають імпульси, які змінюють мембранний потенціал початкового сегмента, тобто є просто виступами тіла клітини, що збільшують площу інтеграції. Якщо дендритне дерево нейрона велике і містить численні пресинаптичні пухирцеподібні закінчення, то виникає змога більшої взаємодії гальмівних і збуджувальних впливів. Потоки імпульсів, що надходять до дендритів і відходять від них, наростають і загасають. Роль дендритів у виникненні електроенцефалограми (ЕЕГ) розглянуто в Розділі 11.
Сучасні дані свідчать, що функція дендритів у забезпеченні нервової діяльності є складнішою. В дендритах можна зафіксувати потенціали дії. В багатьох випадках вони ініційовані в початковому сегменті і переходять ретроградно, проте потенціал дії, що здатний поширюватись, виникає теж і в самих дендритах. Врешті, посилений інтерес виявляється до мікродоменів, що містяться в цитоплазмі багатьох різних типів клітин, і, ймовірно, формування поблизу окремих дендритних шипів пулів Са2+, що збільшує можливість локальних змін в інтенсивності синаптичної провідності й, отже, може стосуватися процесів навчання та пам’яті.
Електрична провідність
У синаптичних контактах, де виникає електрична провідність, імпульс, що надходить до закінчення аксона, генерує в постсинаптичній клітині збуджувальний постсинаптичний потенціал, який унаслідок існування в ділянці контакту низькорезистентних містків має коротший латентний період, ніж ЗПСП у синапсах з хімічною провідністю. В синапсах мішаного типу відбуваються як такого типу коротколатентна реакція, так і довголатентна, опосередкована хімічно.
Виникає внаслідок ушкодження спинного мозку, і у випадку розсіяного склерозу, зокрема, у разі інтратекального введення за допомогою імплантованої помпи. Інші трансмітери теж зумовлюють пресинаптичне гальмування шляхом G-білок-опосередкованої дії на Са2+- і К+ - канали.
І навпаки, у разі пролонгованого потенціалу дії виникає пресинаптичне збудження (див. рис. 4-12), і Са2+ - канали відкриваються на триваліший час. Молекулярні механізми, що забезпечують пресинаптичне збудження і які опосередковують серотоніном, детально вивчені на морських слимаках Aplysia. Виділення в аксо-аксонних закінченнях серотоніну зумовлює збільшення внутрішньоклітинної концентрації циклічного АМФ, і наступне фосфорилювання одного з видів К+ - каналів зумовлює закриття цих каналів, сповільнюючи реполяризацію і пролонгуючи потенціал дії.
Організація гальмівних систем
Пресинаптичне і постсинаптичне гальмування виникають переважно внаслідок збудження в певних системах, що конверґують на конкретному постсинаптичному нейроні (аферентне гальмування). Нейрони можуть теж гальмувати шляхом негативного зворотного зв’язку. Наприклад, кожен мотонейрон спинного мозку регулярно віддає зворотну колатераль, що утворює синаптичний контакт з гальмівним інтернейроном, який закінчується на тілі спинномозкового нейрона, а також інших мотонейронах спинного мозку (рис. 4-13). Цей специфічний гальмівний нейрон інколи називають клітиною Реншоу. Імпульси, що генеруються в мотонейроні, активують гальмівний інтернейрон, який зумовлює виділення гальмівного трансмітера і, отже, сповільнення або навіть припинення розряду в мотонейроні. Подібні гальмування через зворотні колатералі простежуються в корі великого мозку і лімбічній системі.
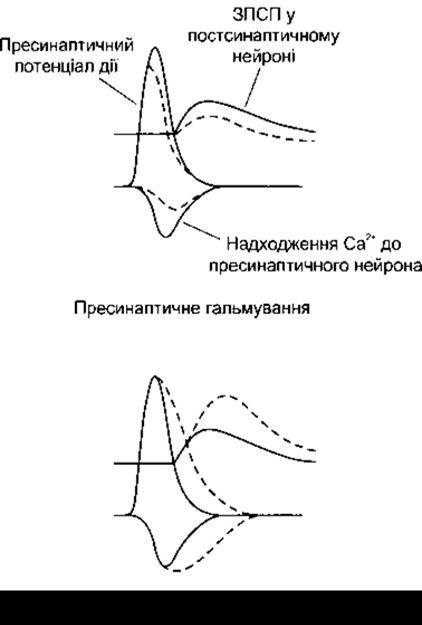
Рис. 4-12. Вплив пресинаптичного гальмування та пресинаптичного полегшення на потенціал дії, надходження Са2+ до пресинаптичного нейрона і ЗПСП в постсинаптичному нейроні. В кожному випадку суцільні лінії відображають контрольні значення, а штрихові - зміни, що виникають унаслідок гальмування і полегшення (модифіковано з Kandel ER, Schwartz JH, Jessell TM [editors]. Principles of Neural Science, 4th ed. McGraw-Hill, 2000).
Пресинаптичне гальмування за участю низхідних шляхів, що закінчуються на аферентних шляхах у задньому розі, може брати участь у передаванні відчуття болю (див. Розділ 7).
Іншого типу гальмування виникає в мозочку. В цій частині головного мозку стимулювання кошикових клітин зумовлює ГПСП у клітинах Пуркіньє (див. Розділ 12). Однак кошикові клітини і клітини Пуркіньє збуджуються тими ж паралельними волокнами. Така організація, яку називають гальмуванням прямим зв’язком, мабуть, обмежує тривалість збудження, зумовленого певними аферентними імпульсами.
Сумація і оклюзія
Взаємодія між збуджувальними і гальмівними впливами в синаптичних контактах нервової мережі відображає інтегрувальну і модулювальну діяльність нервової системи.
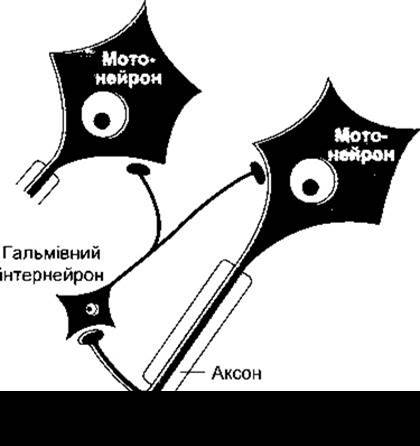
Рис. 4-13. Гальмування зворотним зв’язком спинномозкового мотонейрона за допомогою інтернейрона (клітини Реншоу).
У гіпотетичній нервовій мережі, яка зображена на рис. 4-14, нейрони А і В конверґують на X, а нейрон В диверґує на X і Y. Подразнення А або В зумовлює виникнення ЗПСП в X. Якщо А і В подразнені одночасно і виникають потенціали дії, то в X виникають дві ділянки деполяризації і відбувається сумація їхньої дії. Унаслідок цього ЗПСП в X буде вдвічі більшим, ніж у разі окремого подразнення А або В, і потенціал в X з легкістю досягає критичного рівня деполяризації. Ефект деполяризації, зумовлений імпульсами в А, підсилює імпульси, що спричинюють активність у В, і навпаки; отже, виникає просторове підсилення. В цьому випадку Y не збуджується, проте його збудливість підвищена, і в ньому легше виникне ЗПСП за умови активності нейрона С. Отже, Y, як кажуть, перебуває в підпороговому стані стосовно X.
Загалом нейрони перебувають у підпороговому стані, якщо без впливу аферентних імпульсів (тобто поза зоною розряду), у них підвищена збудливість. Нейрони, що мають на собі декілька активних пухирцеподібних закінчень, перебувають у підпороговому стані, а ці, що мають їх багато - у зоні розряду. Гальмівні імпульси зумовлюють подібне часове та просторове підсилення й ефект підпорогового стану.
Якщо в нейроні В потенціали дії виникають один за одним, то X і Y збуджуються внаслідок часової сумації утворених ЗПСП. Якщо С подразнено декілька разів, то Y і Z теж збуджуються. Якщо В і С збуджуються повторно в цей же час, то розряд буде виникати в X, Y і Z. Отже, відповідь на одночасне подразнення В і С не є більшою, ніж сума відповідей на окремі подразнення В і С, оскільки В і С закінчуються на нейроні Y. Таке зниження рівня очікуваної відповіді, що відбувається в разі подразнення пресинаптичних волокон, називають оклюзією.
Збуджувальний і гальмівний підпорогові ефекти, а також явище оклюзії можуть значно впливати на провідність імпульсів у кожному конкретному шляху. Внаслідок цих впливів часові характеристики під час проходження імпульсів з периферії до головного мозку через низку синапсів змінюються. Ці явища можуть теж пояснювати такий важливий феномен, як передавання відчуття болю (див. Розділ 7).
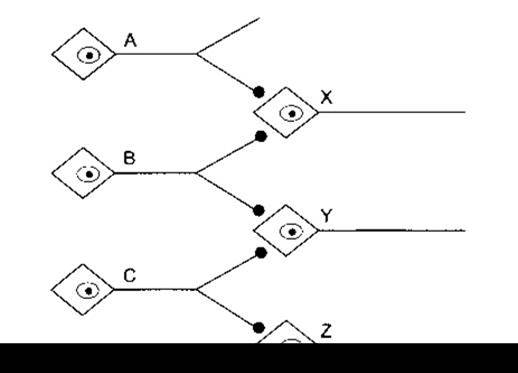
Рис. 4-14. Проста мережа нейронів. Нейрони А, В і С утворюють збуджувальні закінчення на нейронах X, Y і Z.
Нейромодуляція
Термін модуляція у фізіології часто трактують настільки вільно, що його визначення мало інформує про цю функцію. Однак у нейробіології термін нейромодуляція використовують для чіткого означення несинаптичної дії речовин на нейрони, що змінює їхню чутливість до синаптичного збудження або гальмування. Нейромодуляція здебільшого зумовлена дією нейропептидів і циркулювальних стероїдів, а також стероїдів, що продукуються в нервовій системі (нейростероїди; див. нижче).