Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Регулювання постави і рухів
Кірково-спинномозкова і кірково-ядерна система - Структура і функція
Провідні шляхи
Нервові волокна, що йдуть від моторної кори до ядер черепних нервів, формують кірково-ядерний шлях. Нервові волокна, які перетинають серединну лінію в ділянці пірамід довгастого мозку й утворюють латеральний кірково-спинномозковий шлях, становлять 80% кірково-спинномозкового шляху. Решта 20% утворюють передній, або вентральний, кірково-спинномозковий шлях (рис. 12-2), який не перетинає серединної лінії аж доки не сягне рівня, на якому ці волокна формують синаптичні контакти з руховими нейронами. Крім того, цей шлях містить кірково-спинномозкові нейрони, які закінчуються на тому ж боці тіла. Передній шлях, філогенетично старіший, спочатку закінчується на інтернейронах, які утворюють синапси на нейронах медіальної частини переднього рогу, звідки відбувається іннервування мускулатури осьового скелета і проксимальних відділів кінцівок. Відповідно, латеральний кірково-спинномозковий шлях прямує до нейронів латеральної частини переднього рогу, які іннервують мускулатуру дистальних відділів кінцівок і, отже, регулюють тонкі й точні рухи. В людини волокна цієї філогенетично новішої системи безпосередньо закінчуються на латеральних рухових нейронах.
Кіркові моторні поля
Подразнення кіркових полів, з яких починаються кірково-спинномозковий і кірково-ядерний шляхи, спричинює швидкі і диференційовані рухи. Найвідоміша така зона - моторна кора (МІ) передцентральної закрутки (рис. 12- 3). Однак є також додаткове моторне поле, розміщене вище від поясної борозни на медіальній поверхні півкулі, яке сягає передмоторної кори, поширюючись на латеральну поверхню (див. рис. 12-3). Рухові відповіді теж виникають під час подразнення соматосенсорного поля І, розміщеного в зацентральній закрутці, і соматосенсорного поля II, розміщеного з боків Сильвієвої борозни (див. Розділ 7). Ці спостереження узгоджуються з фактом, що 30% волокон кірково-спинномозкового і кірково-ядерного шляхів виходять з моторної кори, 30 - з премоторної, а 40% - з тім’яної частки, зокрема з соматосенсорної ділянки.
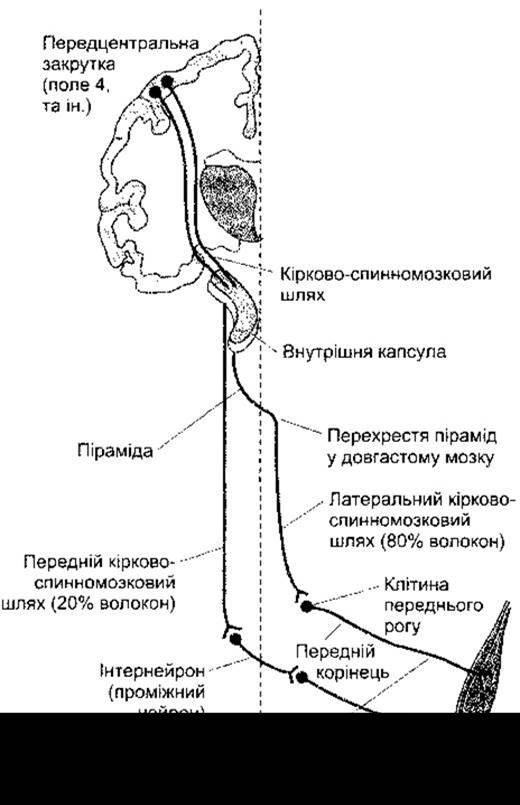
Рис. 12-2. Кірково-спинномозкові шляхи.
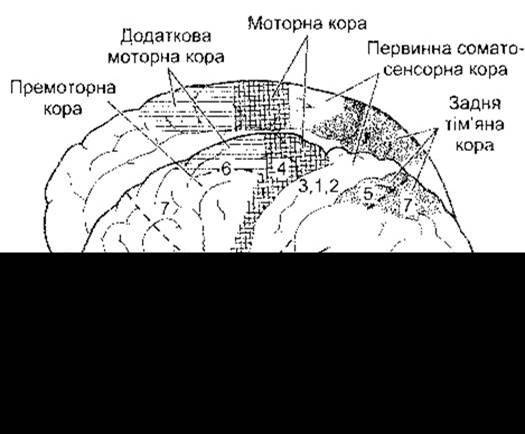
Рис. 12-3. Медіальна (вгорі) і латеральна (внизу) поверхні кори півкуль великого мозку людини. Показано моторну кору (4 поле Бродмана) та інші поля, причетні до регулювання довільних рухів, нумеровані за Бродманом (відтворено за дозволом з Kandel ER, Schwartz JH, Jesseli TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000).
Експериментальне подразнення кори пацієнтів після виконаної під місцевою анестезією трепанації черепа дало змогу ідентифікувати більшість моторних проекцій ділянок кори. Отримані результати підтверджені скануванням ПЕТ і ЯМР головного мозку в людей, які не зазнавали оперативного втручання чи знеболення (рис. 12-4). Різні ділянки тіла відображені в передцентральній закрутці так, що стопа проектується у верхній частині закрутки, а обличчя - внизу (рис. 12-5). Поле обличчя відтворене білатерально (в обидвох півкулях), тоді як решта тіла - унілатерально і таким способом, що кіркове моторне поле регулює роботу мускулатури протилежної частини тіла. Відображення в корі кожної з ділянок тіла за розмірам пропорційне до майстерності виконання довільних рухів. Наприклад, ділянки кори, пов’язані з регулюванням артикуляції мови чи рухами, що їх виконують м’язи кисті, є особливо широкими; використання гортані, губ і язика для вимови слів, а також пальців, особливо великого, забезпечує діяльність, що досягає в людини найвищого рівня. Умови, у яких можливе дослідження кори великого мозку людини, не дають змоги подразнювати краї борозен та інші недоступні ділянки. Детальними дослідженнями на мавпах виявлено, що в корі є відповідне відображення частин тіла, причому осьова мускулатура і проксимальні ділянки кінцівок відображені вздовж переднього краю передцентральної закрутки, а дистальні ділянки кінцівок - уздовж її заднього краю. Іншою особливістю МІ є як наявність значного перекриття ділянок іннервації м’язів, так і значна розмежованість зон іннервації м’язів-синергістів. Тривають дискусії щодо питання: у МІ проектуються окремі м’язи чи певні рухові реакції; найновіші ж дані свідчать, що в ній проектуються як ті, так і інші. Клітини ділянок моторної кори впорядковані у вигляді окремих колонок, або стовпців. Клітини кожного стовпця отримують численні чутливі вхідні імпульси з периферійної ділянки, у якій вони регулюють рухи, і таким способом відбувається забезпечення основи для контролю рухів за зворотним зв’язком. Деякі з цих імпульсів є прямими, а інші перемикаються в сенсорному полі І зацентральної закрутки.
Рис. 12-4. Ділянка кисті моторної кори семирічного хлопчика, зображена за допомогою ЯМР. Зміни активності, зумовлені стисканням гумового м’ячика правою кистю, відображені білим кольором, лівою кистю - чорним кольором. У цьому окремому випадку активування лівої рухової кори руховою активністю лівої руки (див. текст) не помітне (відтворено за дозволом з Waxman SG: Correlative Neuroanatomy, 24th ed. McGraw-Hill, 2000).
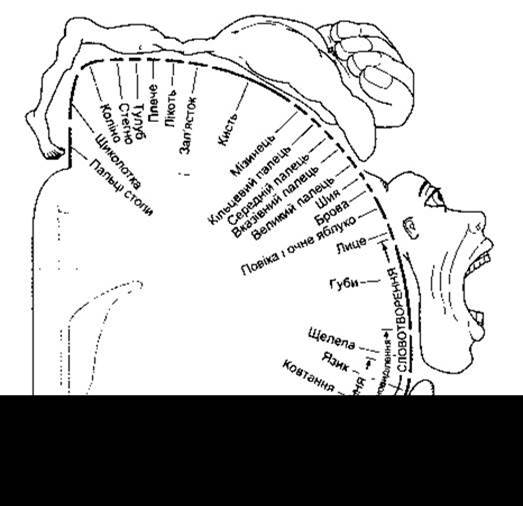
Рис. 12-5. Руховий гомункул. Схема фронтального перерізу передцентральної закрутки, що відображає кіркове відображення різних ділянок тіла. Розміри різноманітних ділянок тіла гомункула пропорційні до відповідних кіркових полів закрутки (відтворено за дозволом з Penfield W, Rasmussen G: The Cerebral Cortex of Man. Macmillan, 1950).
Домінування головного мозку, про що детально йдеться в Розділі 16, стосується і моторної кори людини. Рухи пальців лівої кисті переважно супроводжуються активуванням правої моторної кори, і навпаки, як досліджено за допомогою ЯМР (див. Розділ 32); проте рухи пальців лівої кисті теж активують і ліву моторну кору, зокрема в осіб, які ліпше володіють правою рукою. Це узгоджується з фактом, що ушкодження лівої моторної кори спричинює порушення функції лівої кисті, як і правої, тоді як у разі ушкодження правої моторної кори функція правої кисті послаблюється незначно.
Пластичність
Завдяки ПЕТ і функційному ЯМР з’ясовано, що в інтактних експериментальних тварин, як і в людини, моторна кора має таку ж пластичність, як і сенсорна (див. Розділ 7). Тому, наприклад, ділянка відображення пальців у моторній корі контрлатерального боку збільшується в розмірах завдяки тренуванню швидких рухів пальців однієї руки; ці зміни помітні вже через тиждень і досягають максимального рівня на четвертий тиждень. Ділянки кори, що регулюють скорочення інших м’язів, також збільшуються в розмірах унаслідок тренування. Незважаючи на невелике ішемічне ушкодження ділянки відображення кисті в моторній корі мавпи, з відновленням рухової функції відновлюється ділянка відображення кисті в сусідній, не ушкодженій частині кори. Отже, карта відображення в моторній корі змінюється з набуттям досвіду.
Додаткове моторне поле
Більшість імпульсів з додаткового моторного поля прямує до моторної кори. Відповідно, це поле бере участь у програмуванні рухових послідовностей. Ушкодження його в мавп спричинює виникнення труднощів у виконанні складних комбінованих рухів і порушення бімануальної координації.
Якщо людина підраховує мовчки, то моторна кора бездіяльна, проте якщо вона починає вимовляти цифри, якими маніпулює, то притік крові до моторної кори і додаткового моторного поля зростає. Отже, додаткове моторне поле разом з моторною корою забезпечують складні довільні рухи, пов’язані з їхнім попереднім плануванням. Притік крові зростає незалежно від того, виконується планований рух чи ні. Збільшення притоку крові простежується в будь-якому випадку, тобто незважаючи на те, чи планований рух виконує контрлатеральна кисть, чи іпсилатеральна.
Премоторна кора
Імпульси з премоторної кори надходять до ділянок стовбура головного мозку, причетних до регулювання постави, і до моторної кори, зокрема до її ділянок, з яких починаються кірково-спинномозковий і кірково-ядерний шляхи. Функція премоторної кори досі остаточно не з’ясована; вважають, що вона пов’язана з регулюванням постави на початку планованого руху, зокрема з підготовкою до його виконання.
Задня тім’яна кора
Крім волокон, що є в складі кірково-спинномозкового і кірково-ядерного шляхів, з соматосенсорного поля і прилеглої ззаду ділянки тім’яної частки відходять волокна до премоторної кори. Ушкодження соматосенсорного поля спричинює дефекти у виконанні завчених послідовностей рухів, наприклад, користування під час їди ножем і виделкою. Частина нейронів поля 5 (див. рис. 12-3) пов’язана з регулюванням таких рухів, як націлювання кисті в напрямі будь-якого об’єкта або маніпулювання нею, а деякі нейрони поля 7 причетні до координування реакції кисть-око.
Роль у регулюванні рухів
Кірково-спинномозковий і кірково-ядерний шляхи, як зазначено, - це система забезпечення тонких і точних рухів. Однак це не означає, що рухи, навіть тонкі, неможливі без участі цієї системи. Нижчі хребетні, по суті, не мають кірково-спинномозкової і кірково-ядерної системи, проте вони досить спритно пересуваються. Коти і собаки можуть стояти, ходити, бігати і навіть їсти після цілковитого руйнування цієї системи. Лише в приматів таке втручання спричинює суттєві зміни.
Обережне перетинання пірамід, що зумовлює високо-вибіркову деструкцію латерального кірково-спинномозкового шляху в лабораторних приматів, призводить до раптової і тривалої втрати здатності до захоплення й утримування малих предметів двома пальцями та ізольованих рухів у зап’ястку. Ці порушення зумовлені втратою регулювання дистальної мускулатури кінцівок, що в нормі забезпечує тонкі і точні рухи. З іншого боку, ушкодження переднього кірково-спинномозкового шляху спричинює порушення функцій осьових м’язів з затруднениям підтримки рівноваги, ходьби і підіймання вгору.
Вплив на рефлекси розтягування
Перетинання пірамід у мавпи зумовлює переважно тривалу гіпотонію і млявість, а не еластичність. У людини, з огляду на особливості будови відділів ЦНС, патологічний процес лише зрідка ушкоджує кірково-спинномозковий і кірково-ядерний шляхи ізольовано, тобто без ушкодження шляхів, що беруть участь у регулюванні постави. Поява еластичності свідчить, очевидно, про ушкодження власне цих шляхів, а не кірково-спинномозкового і кірково-ядерного.
У випадку ушкодження латерального кірково-спинномозкового шляху в людини виникає симптом Бабінського: тильне згинання великого пальця і віялоподібне положення решти пальців стопи під час подразнення шкіри латерального краю підошви. За винятком немовлят, нормальним реагуванням на це подразнення є підошовне згинання всіх пальців стопи. Симптом Бабінського означає зникнення цього згинального рефлексу, що в нормі використовують для оцінки функції латерального кірково-спинномозкового шляху. Він є важливий для визначення локалізації патологічного процесу, однак фізіологічні підстави його невідомі.