Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Регулювання постави і рухів
Мозочок
Структурний поділ
Мозочок розміщений над важливими чутливими і руховими системами стовбура головного мозку (рис. 12-13). Він сполучений зі стовбуром головного мозку з кожного боку верхніми мозочковими ніжками, середніми, або мостовими, мозочковими ніжками і нижніми мозочковими ніжками. Середня частина мозочка - черв’як - і розміщені латерально півкулі мозочка набагато складчастіші і мають глибші борозни, ніж кора великого мозку; маса мозочка становить лише 10% від маси кори великого мозку, проте площа його поверхні - близько 75% від площі кори великого мозку. Анатомічно мозочок розділений двома поперечними щілинами на три частини. Задньолатеральна щілина відмежовує розміщений медіально вузлик і розміщений латерально жмуток з кожного боку від решти мозочка, а перша щілина розділяє решту мозочка на передню і задню частки. Менші щілини розділяють черв’як на десять первинних часточок, які нумерують зверху донизу від 1 до X. Ці часточки зображені на рис. 12-14.
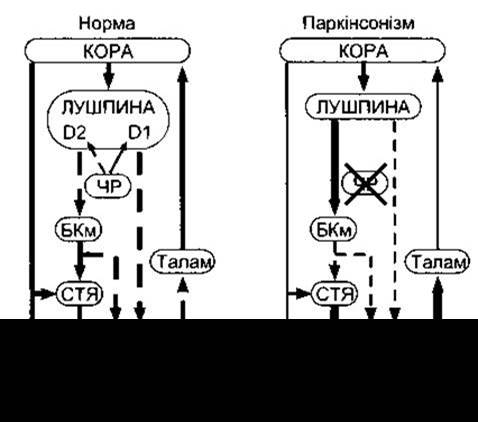
Рис. 12-12. Базальноядерно-таламокортикальний цикл у випадку хвороби Паркінсона. Суцільні стрілки означають збуджувальні, а штрихові - інгібувальні впливи. Сила кожного впливу означена різної товщини стрілками. Смугасте тіло містить D1-дофамінові рецептори, що підсилюють безпосередній вплив на медіальну бліду кулю (БКм), і D2-рецептори, що знижують рівень непрямого інгібувальйого впливу на БКм через лушпину, латеральну бліду кулю (БКл) і субталамічні ядра (СТЯ). ЧP - чорна речовина; Талам - таламус; НМЯ - ніжко-мостові ядра (відтворено за дозволом з Grafton SC, De-Long М: Tracing the brain’s circuitry with functional imaging. Nat Med 1997;3:602).
Функціональний поділ
З погляду функцій мозочок теж поділяють на три частини, проте цей поділ відрізняється від анатомічного (рис. 12-15). Вузлик черв’яка і жмуток півкулі кожного боку утворюють жмутково-вузликову частку, або вестибулярний (присінковий) мозочок. Ця частка, що є філогенетично найстаршою частиною мозочка, пов’язана з вестибулярною системою і важлива для підтримання рівноваги та зумовлених навчанням змін вестибулоочного рефлексу (див. Розділ 9). Решта черв’яка і суміжні з ним медіальні ділянки півкуль становлять спінальний мозочок, до якого проходять пропріорецептивні шляхи від тіла, а також шляхи, по яких надходить інформація щодо відображення частин тіла в моторній корі. Зіставляючи планування з реалізацією виконання рухів у частинах тіла, мозочок забезпечує плавність і координованість рухів. Зв’язки черв’яка мозочка з ділянками стовбура головного мозку потрібні для регулювання осьових м’язів і м’язів проксимальних відділів кінцівок, тоді як такі ж зв’язки півкуль мозочка зі стовбуром головного мозку - для регулювання м’язів дистальних відділів і кінцівок. Латеральні ділянки півкуль мозочка називають новим мозочком. Ці з філогенетичного погляду найновіші структури мозочка досягають найбільшого розвитку в людини. Разом з моторною корою структури нового мозочка виконують планування і програмування рухів.
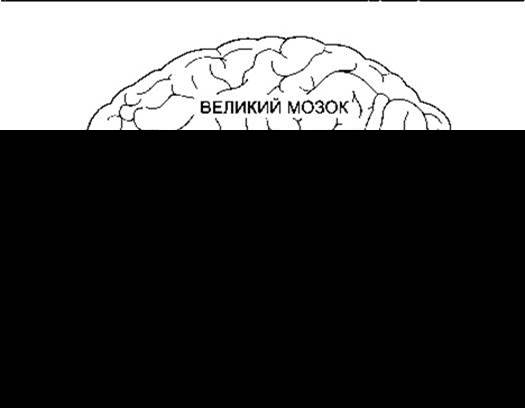
Рис. 12-13. Головні частини головного мозку. Деяке зміщення частин на рисунку дає змогу показати мозочкові ніжки, а також структуру у формі кільця, утворену мозочком, передньою частиною мосту і середньою мозочковою ніжкою, що «пеленає» стовбур головного мозку (відтворено за дозволом з Goss CM [editor]: Gray’s Anatomy of the Yuman Body, 27th ed. Lea & Febiger, 1959).
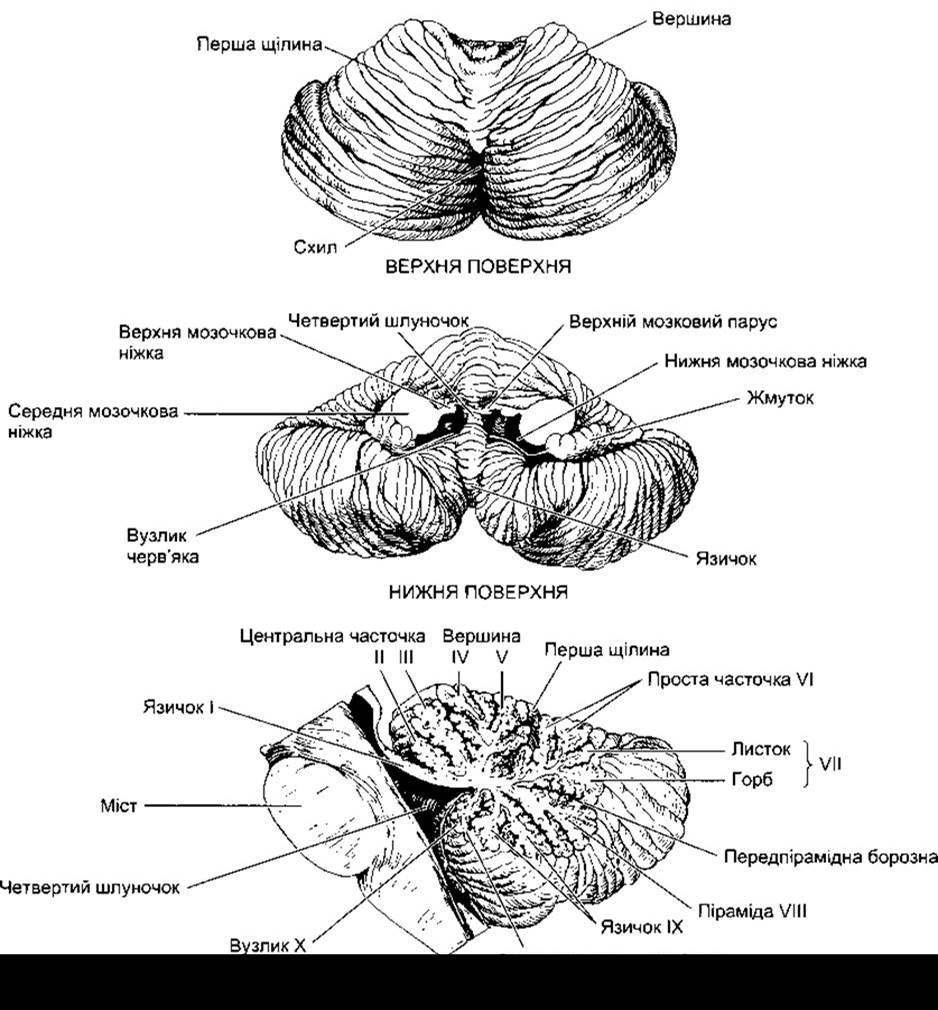
Рис. 12-14. Верхня та нижня поверхні і сагітальний переріз мозочка людини. Позначено (від І до X) і наведено назви головних часточок черв’яка мозочка.
Організація мозочка
Мозочок складається з розміщеної ззовні кори мозочка, білої речовини і розташованих глибоко в ній ядер мозочка. Первинні аферентні шляхи мозочка, мохоподібні і ліаноподібні волокна (див. нижче) утворюють колатералі з глибокими ядрами і надходять до кори (рис. 12-16). Розрізняють чотири ядра мозочка: зубчасте, кулясте, кіркоподібне і ядро шатра.
Кулясте і кіркоподібне ядра інколи
об’єднують назвою проміжне ядро. Більша частина шляхів від вестибулярного мозочка проходить безпосередньо до стовбура головного мозку, а решта - від кори мозочка до його ядер, звідси - до стовбура головного мозку. Отже, ядра мозочка забезпечують єдиний шлях імпульсів від спінального і нового мозочків. Медіальна частина шляху від спінального мозочка проходить до ядра шатра, а звідси - до стовбура головного мозку. Шлях від суміжних частин півкулі мозочка, що теж належать до спінального мозочка, проходить до кіркоподібного та кулястого ядер, а від них - до стовбура головного мозку. Волокна від нового мозочка йдуть до зубчастого ядра, а від нього безпосередньо або опосередковано - до вентролатерального ядра таламуса.
Рис. 12-15. Поділ мозочка за функціями (модифіковано з Kandel ER, Schwartz JH, Jesseli TM [editors]: principles of Neural Science, 4th ed. McGraw-Hill, 2000)
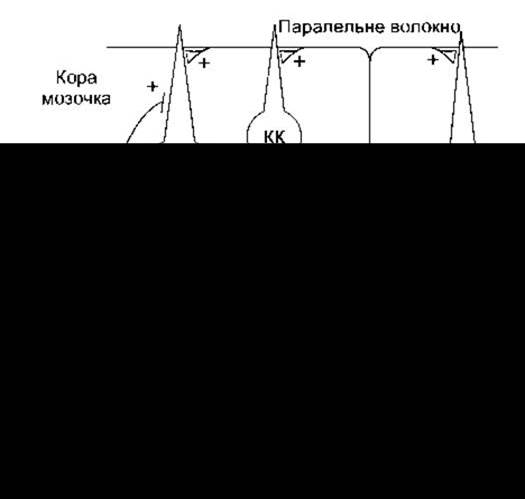
Рис. 12-16. Схема нервових зв’язків усередині мозочка. Плюс і мінус позначають, відповідно, збуджувальні і гальмівні впливи. КК - кошикова клітина; КҐ - клітина Ґольджі; ЗК - зерноподібна клітина, КЯМ - клітина ядра мозочка; КП - клітина Пуркіньє. Зверніть увагу на те, що КП і КК чинять гальмівний вплив. Зв’язки зірчастих клітин подібні до зв’язків кошикових клітин, проте в більшості вони закінчуються на дендритах клітин Пуркіньє
Кора мозочка містить п’ять різновидів нейронів: клітини Пуркіньє, зерноподібні клітини, кошикові клітини, зірчасті клітини і клітини Ґольджі. Вона має тришарову структуру (рис. 12-17): зовнішній, молекулярний шар, гангліонарний шар, утворений одним шаром клітин Пуркіньє, і внутрішній, зернистий, шар. Клітини Пуркіньє - одні з найбільших нейронів тіла. їхні значно розгалужені дендрити проходять крізь молекулярний шар кори. Аксони клітин Пуркіньє, що формують єдиний еферентний шлях від кори мозочка, йдуть переважно до ядер мозочка. Кора мозочка теж містить зерноподібні клітини, які отримують імпульси через мохоподібні волокна і передають їх на клітини Пуркіньє. Тіла зернистих клітин розміщені в зернистому шарі. Аксон кожної зернистої клітини проникає в молекулярний шар, де утворює Т-подібне розгалуження. Ці довгі прямі розгалуження названо паралельними волокнами. Дендрити клітин Пуркіньє утворюють численні кущоподібні розгалуження, розміщені в одній площині й орієнтовані під прямими кутами стосовно паралельних волокон. Паралельні волокна створюють синаптичні контакти з дендритами багатьох клітин Пуркіньє і, отже, разом з дендритами клітин Пуркіньє формують дивовижну регулярну сітку.
Інші три різновиди нейронів кори мозочка є інгібіторними інтернейронами. Кошикові клітини (див. рис. 12- 17) розміщені в молекулярному шарі. Вони отримують імпульси від паралельних волокон, і кожна кошикова клітина передає інформацію багатьом клітинам Пуркіньє. Аксони клітин утворюють розгалуження у формі кошика навколо тіла і горбка аксона клітин Пуркіньє. Зірчасті клітини подібні до кошикових, проте розташовані ближче до поверхні кори. Клітини Гольджі розміщені в зернистому шарі. їхні дендрити, що проникають у молекулярний шар, одержують інформацію від паралельних волокон. Тіла клітин отримують імпульси через колатералі від мохоподібних волокон і клітин Пуркіньє. Аксони клітин Ґольджі контактують з дендритами зернистих клітин.
Як зазначено вище, головні шляхи збуджувальних впливів, що надходять до кори мозочка, сформовані ліано- і мохоподібними волокнами (див. рис. 12-16). Ліаноподібні волокна походять з єдиного джерела - ядер оливи - і проходять до первинних дендритів клітин Пуркіньє та звиваються навколо них як ліани. До ядер оливи пропріорецептивні імпульси надходять від усіх частин тіла. З іншого боку, мохоподібні волокна передають інформацію безпосередньо від усіх ділянок тіла, а також імпульси від кори великого мозку через ядра мосту до кори мозочка. Ці волокна закінчуються на дендритах зернистих клітин складними синаптичними скупченнями, які називають клубочками. Ці клубочки містять також терміналі клітин Ґольджі, що чинять гальмівний вплив, про які згадувалось раніше.
Головні цикли циркуляції збудження в корі мозочка порівняно нескладні (див. рис. 12-16). Імпульси, які передає ліаноподібне волокно, чинять потужний збуджувальний вплив на поодинокі клітини Пуркіньє, тоді як мохоподібні через зернисті клітини забезпечують слабкий збуджувальний вплив на низку клітин Пуркіньє. Кошикові і зірчасті клітини збуджувані зернистими клітинами через паралельні волокна і їхній вплив на клітини Пуркіньє є гальмівним (гальмування прямим зв’язком). Клітини Ґольджі збуджують імпульси, що надходять через колатералі мохоподібних волокон, колатералі клітин Пуркіньє та паралельні волокна і гальмують передавання імпульсів від мохоподібних волокон до зернистих клітин.
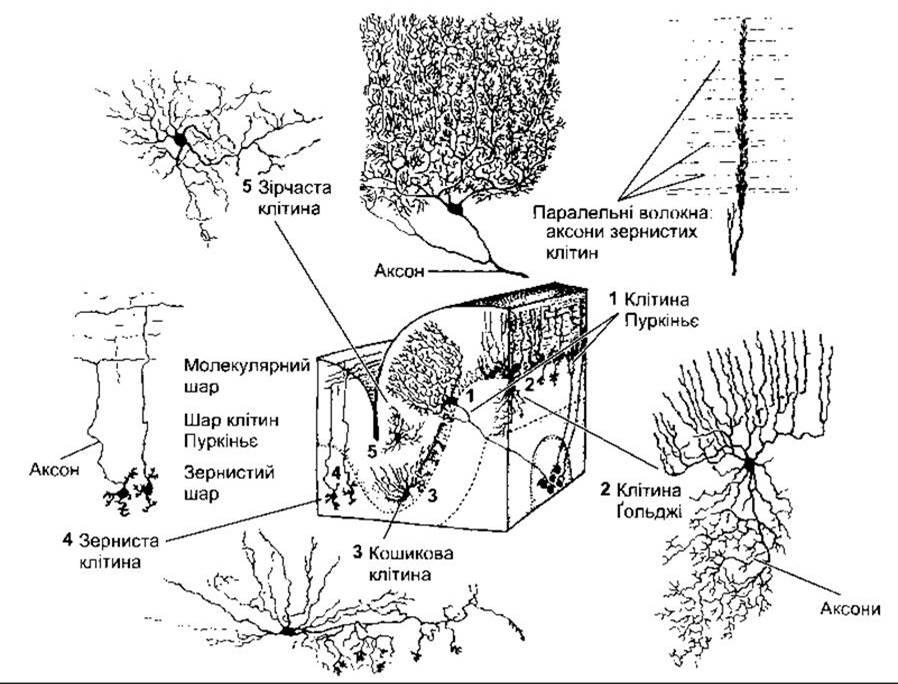
Рис. 12-17. Розміщення і структура нейронів кори мозочка (відтворено за дозволом з Kuffler SW, Nicholls JG, Martin AR: From Neuron to Brain, 2nd ed. Sinauer, 1984).
Нейротрансмітером, що його, ймовірно, виділяють зірчасті, кошикові клітини, клітини Ґольджі та клітини Пуркіньє, є ГАМК, тоді як зернисті клітини виділяють глютамат. ГАМК діє на ГАМКА-рецептори, проте комбінація субодиниць у цих рецепторах (див. Розділ 4) в різних типів клітин різна. Унікальність зернистих клітин полягає в тому, що вони - це єдиний різновид нейронів ЦНС, ГАМКА-рецептор яких містить а6-субодиницю.
Імпульси, що надходять з клітин Пуркіньє, відповідно, чинять гальмівний вплив на ядра мозочка. Як зазначено вище, ці ядра теж отримують збуджувальні імпульси через колатералі ліано- і мохоподібних волокон. Цікаво, що, незважаючи на гальмівний вплив клітин Пуркіньє на ядра мозочка, імпульси, які надходять від цих ядер до стовбура головного мозку і таламуса, є збуджувальними. Отже, майже всі шляхи циркуляції збудження в мозочку, очевидно, стосуються модуляції чи регулювання синхронності передавання імпульсів від ядер мозочка до стовбура головного мозку і таламуса.
Первинні аферентні шляхи, що проходять до мозочка мохо- і ліаноподібними волокнами, наведені в табл. 12-5.
Жмутково-вузликова частка
Тварини, у яких зруйнована жмутково-вузликова частка, втрачають стійкість і пересуваються, хитаючись на рівній площині. Вони не підтримують рівноваги і можуть впасти без сторонньої підтримки. Подібні порушення, що є ранніми ознаками розвитку пухлини в серединних ділянках мозочка, яка походить з клітин вузлика, можна спостерігати в дітей. У цьому разі швидко ушкоджуються структури, розміщені в межах жмутково-вузликової частки.
Вибіркове хірургічне видалення жмутково-вузликової частки в собак спричинює синдром хвороби руху (див. Розділ 9), тоді як широке ушкодження інших ділянок мозочка чи головного мозку не має такого наслідку.
Вплив на рефлекси розтягування
Подразнення ділянок мозочка, до яких надходять пропріорецептивні імпульси, в деяких випадках гальмує, а в деяких - підсилює рухи, спричинені подразненням кори великого мозку. Ушкодження листків I-VI та присерединних ділянок зумовлює в експериментальних тварин еластичність у частинах тіла, що проектовані в ушкоджених ділянках мозочка. Однак у людей відповідні деструктивні зміни в мозочку призводять до гіпотонії м’язів.
Вплив на рухи
За винятком змін, що простежуються в рефлексах розтягування, в експериментальних тварин і людей з ушкодженням півкуль мозочка в стані спокою не зафіксовано інших відхилень. Проте під час рухів починають виявлятись різко виражені ненормальності. Ці вади ані не є паралічем, ані не пов’язані з втратою чутливості, а мають характер виразної атаксії, що виявляється в дискоординації, зумовленій порушенням швидкості, амплітуди, сили і напряму рухів. У цьому разі атаксія може виявлятись на одній половині тіла. У випадку ушкодження лише кори мозочка вади руху внаслідок компенсації поступово зникають. Ушкодження ядер мозочка спричинює вираженіші та стійкіші порушення. З огляду на це в разі хірургічного втручання на мозочку треба уникати ушкодження його ядер.
Таблиця 12-5. Функція головних аферентних систем мозочка1
Аферентні шляхи |
Проводять |
Присінково-мозочковий |
Вестибулярні імпульси з лабіринтів безпосередньо або через присінкові ядра |
Задній спинномозочковий |
Пропріо- й екстерорецептивні імпульси від тіла |
Передній спинномозочковий |
Пропріо- й екстерорецептивні імпульси від тіла |
Клиномозочковий (дорсальна частина зовнішніх дугоподібних пучків) |
Пропріорецептивні імпульси, зокрема від ділянок голови та шиї |
Покришко-мозочковий |
Слухові і зорові імпульси, що проходять через нижні і верхні горбики покришки середнього мозку |
Мосто-мозочковий |
Імпульси від моторної та інших ділянок кори великого мозку, що йдуть через ядра моста |
Оливо-мозочковий |
Пропріорецептивні імпульси від усього тіла, що йдуть через ядра оливи |
1 Оливо-мозочковий шлях проходить до кори мозочка ліаноподібними волокнами, інші перечислені шляхи - мохоподібними волокнами. Ще деякі шляхи проводять імпульси від ядер стовбура головного мозку до кори і ядер мозочка, зокрема серотонінергічний шлях від ядер шва до молекулярного і зернистого шарів, а також норадренергічний шлях від голубої плями до всіх трьох шарів.
Ще інші ознаки ушкодження мозочка в людини свідчать про важливість цієї частини мозку для регулювання рухової діяльності. Атаксія виявляється не тільки, грубо кажучи, хиткою, «п’яною» ходою, а також порушенням тонких і точних рухів, що особливо позначається на процесі мовлення. Мова стає невиразною або, як її називають, - скандованою. Інші довільні рухи є теж значно ненормальними. Наприклад, у разі намагання доторкнутися до певного об’єкта палець опиняється то з одного, то з іншого боку від цього об’єкта. Така дисметрія швидко ініціює намагання корекції руху, проте внаслідок грубої корекції палець знову опиняється з іншого боку предмета і так осцилює туди й назад. Подібного типу коливання в разі ушкодження мозочка називають тремором наміру. На відміну від тремору у випадку паркінсонізму, що наявний у стані спокою, тремор наміру виникає тоді, коли пацієнт намагається виконати певний довільний рух. Іншим прявом ушкодження мозочка є нездатність «швидко гальмувати», тобто раптово припиняти виконуваний рух. У нормі, наприклад, під час згинання передпліччя за умов долання певного опору можна швидко припинити рух, якщо цей опір раптом зникає. Хворий з ушкодженням мозочка не може припинити рух кінцівки і його передпліччя рухається назад, описуючи широку дугу. Таку ненормальну реакцію називають феноменом рикошета, такого типу вади виявляються і в разі виконання інших рухів. Це є однією з важливих причин прояву в таких пацієнтів адіадохокінезії - неможливості послідовно виконувати протилежні за напрямом рухи, такі як, наприклад, послідовно повторювальні пронація і супінація кисті. Пацієнти з ушкодженням мозочка мають також утруднення з виконанням рухів одночасно в декількох суглобах. Ці рухи в них відбуваються роздільно в часі в кожному з суглобів. Такий феномен відомий за назвою декомпозиція рухів.
Мозочок і навчання
Функція мозочка пов’язана з пристосуванням за допомогою навчання, і коли завдання виконують раз за разом, то координація рухів поліпшується. Під час виконання рухового завдання активність у головному мозку переміщується з передлобової ділянки до кори тім’яної частки, моторної кори і мозочка. Ймовірно, основу навчання в мозочку становлять імпульси, що надходять з ядер оливи. Щодо цього зазначимо, що кожна клітина Пуркіньє отримує імпульси аж від 250 000 до 1 млн мохоподібних волокон, проте лише від одного ліаноподібного волокна, що проходить від оливи. Це волокно утворює 2 000-3 000 синаптичних контактів з клітиною Пуркіньє. Активування ліаноподібного волокна спричинює в клітині Пуркіньє сильний складний пік активності, і цей пік певним чином зумовлює тривалу модифікацію імпульсів, що через мохоподібні волокна надходять до клітини Пуркіньє. Активність у ліаноподібних волокнах зростає з кожним новим навчальним рухом, а вибіркове ушкодження оливного комплексу спричинює нездатність до формування тривалих пристосувальних механізмів, необхідних для виконання певних рухових відповідей. Роль мозочка в регулюванні вестибулоочного рефлексу й інших форм рефлекторної пам’яті розглянуто в Розділі 16.
Механізми
Хоча функції жмутково-вузликової частки, спінального мозочка і нового мозочка порівняно зрозумілі, а мозочкові цикли не такі складні, проте точні механізми дії його частин досі невідомі. Співвідношення електричних явищ у мозочку з дією його як центру рухової діяльності - це одна з цікавих нез’ясованих проблем. Корі мозочка властивий базальний, 150-300/с, 200 мкВ, електричний ритм, на який накладається 1000—2000/с компонент з меншою амплітудою. Отже, частота базального ритму більше ніж у 10 разів вища від зареєстрованого таким самим способом кіркового a-ритму мозочка. Імпульси, що надходять до мозочка, звичайно змінюють амплітуду його ритму подібно до того, як радіомовний сигнал модулює несучу частоту радіопередавача. Однак значимість цього біоелектричного феномену для функції мозочка не з’ясована.