Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Основы молекулярной биотехнологии
ДНК, РНК и синтез белка
Регуляция транскрипции у бактерии
Все процессы, протекающие в бактериальной клетке, — образование аминокислот, нуклеотидов и других важных метаболитов, репликация, транскрипция, трансляция, катаболизм, высвобождение энергии, реакции на внешние воздействия — требуют участия белков. Однако энергетических ресурсов клетки не хватает для одновременного осуществления транскрипции и трансляции (экспрессии) всех структурных генов. Поэтому постоянно экспрессируются только те гены, которые кодируют белки, поддерживающие основные клеточные функции, а транскрипция остальных структурных генов регулируется. Когда у клетки возникает потребность в каком-то белке (белках), то инициируется (включается) транскрипция соответствующего структурного гена (генов), а когда такая потребность исчезает, транскрипция выключается.
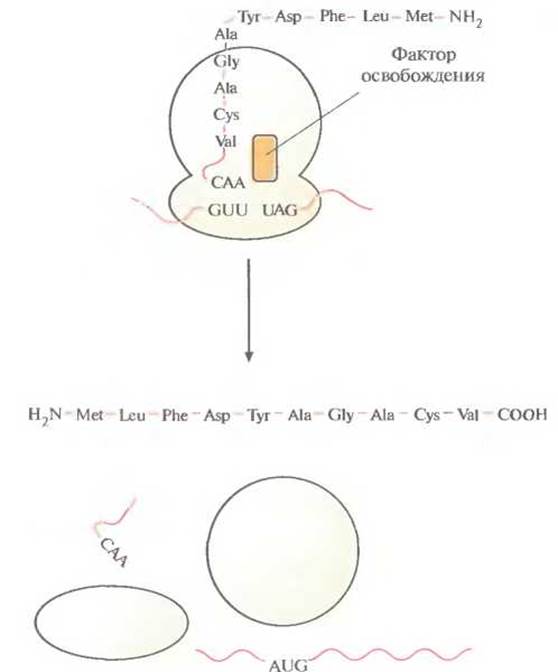
Рис. 3.18. Терминация трансляции. Со стоп-кодоном (UAG) связывается фактор освобождения, и трансляция завершается. Химическая связь между последней тРНК и полипептидной цепью разрывается, свободная тРНК, мРНК и готовая белковая цепь отсоединяются от рибосомы, и последняя диссоциирует на субъединицы.
Часто у бактерий белки одного метаболического пути кодируются смежными структурными генами. Нуклеотидная последовательность, в которой закодировано более одного белка, называется опероном. Обычно оперон находится под контролем единственного промотора, и при его транскрипции образуется одна длинная молекула мРНК, кодирующая несколько белков. При трансляции такой мРНК, в которой стоп-кодон последовательности, кодирующей один белок, соседствует со старт-кодоном гена следующего белка, синтезируется набор дискретных белков.
В большинстве структурных генов Е. coli имеются два сайта связывания для РНК-полимеразы. Один из них обычно представляет собой нуклеотидную последовательность
ТАТААТ
АТАТТА
(ТАТА-бокс, или бокс Прибнова), а другой —
TTGAC
AACTG.
ТАТА-бокс и последовательность TTGAC расположены за 10 (область-10) и 35 (область -35) нуклеотидов до сайта инициации транскрипции соответственно (нуклеотид +1) (рис. 3.19). Обычно от участка между ТАТА-боксом и нуклеотидом +1 во многом зависит, будет ли происходить транскрипция данного оперона. В зависимости от способа регуляции транскрипции оперона этот участок называется оператором или активатором.
Для включения и выключения разных оперонов в ходе эволюции сформировалось множество регуляторных систем. Например, с операторной областью может быть связан регуляторный белок, называемый репрессором; он мешает перемещению РНК-полимеразы вдоль молекулы ДНК, и транскрипция блокируется (рис. 3.20). Однако если с репрессором свяжется некое низкомолекулярное вещество (эффектор), то его конформация изменится таким образом, что его связывание с операторной областью станет невозможным, и транскрипция возобновится. Обычно эффектор разрушается клеточными ферментами. Когда его концентрация снижается, репрессор связывается с операторным участком, и транскрипция вновь прекращается. Операторный участок специфичен для каждого оперона, а эффектор взаимодействует только с определенным репрессором. В качестве иллюстрации рассмотрим такой пример. Предположим, что клетка способна метаболизировать определенный сахар. Тогда синтез ферментов, расщепляющих этот сахар, будет бесполезной тратой клеточных ресурсов, если он отсутствует в среде. С другой стороны, если этот сахар имеется в достаточном количестве и является единственным источником углерода, то ферменты, отвечающие за его утилизацию клеткой, становятся совершенно необходимыми. В этом случае сахар действует как эффектор, препятствуя связыванию репрессора с операторным участком и таким образом обеспечивая транскрипцию оперона и синтез ферментов.
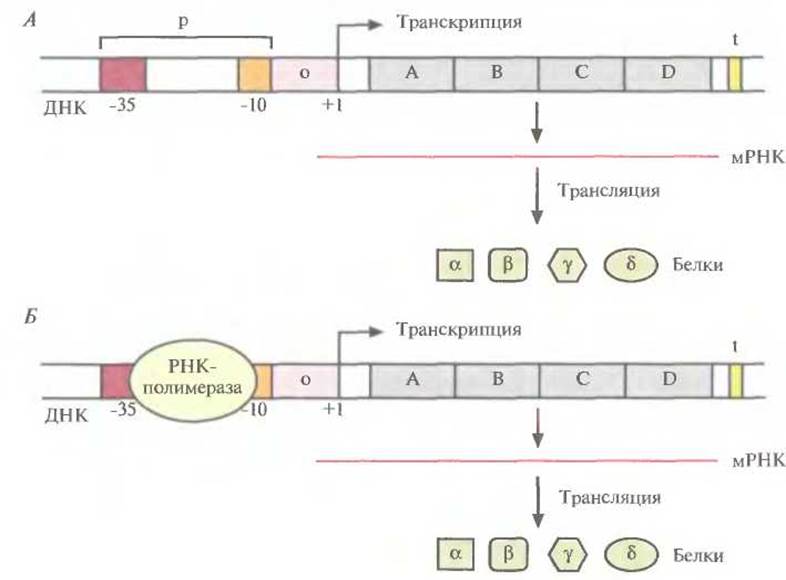
Рис. 3.19. Транскрипция в бактериальной клетке. А. Структурные гены (А, В, С и D) оперона находятся под транскрипционным контролем оператора (о) и промотора (р). РНК-полимераза связывается с участками, находящимися на расстоянии 10 (-10) и 35 (-35) пар оснований от сайта инициации транскрипции (+1). t — Стоп-сигнал, ответственный за остановку транскрипции, а, ß, у и δ — белки, продукты генов А, В, С, D. Б. То же, что и на рис. А. но показано связывание РНК-полимеразы с промоторной областью.
При истощении запасов сахара в среде репрессор связывается с операторным участком, и транскрипция оперона прекращается. Нормальным состоянием других оперонов может быть состояние, при котором осуществляется их транскрипция, поскольку репрессорный белок неактивен. В этом случае специфический эффектор (корепрессор), связываясь с неактивным репрессором, вызывает в нем такие конформационные изменения, которые обеспечивают связывание комплекса с операторным участком, и транскрипция оперона выключается (рис. 3.21). Сам по себе репрессор не способен связываться с оператором, поэтому при уменьшении концентрации корепрессора транскрипция возобновляется.
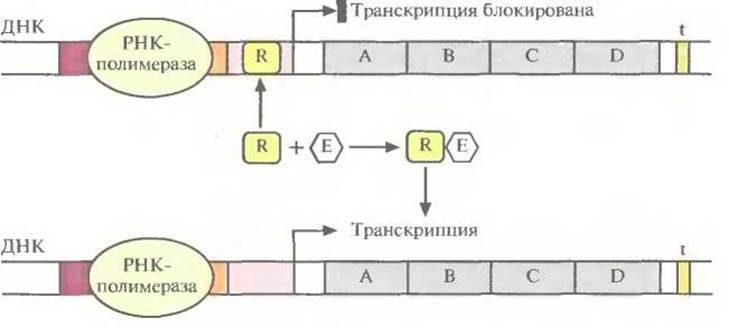
Рис. 3.20. Включение транскрипции бактериального оперона. Репрессор (R) связывается с оператором и блокирует транскрипцию. Связывание эффектора (Е) с репрессором изменяет его конформацию, и он не может связаться с оператором РНК-полимераза беспрепятственно перемещается вдоль молекулы ДНК, осуществляя транскрипцию.
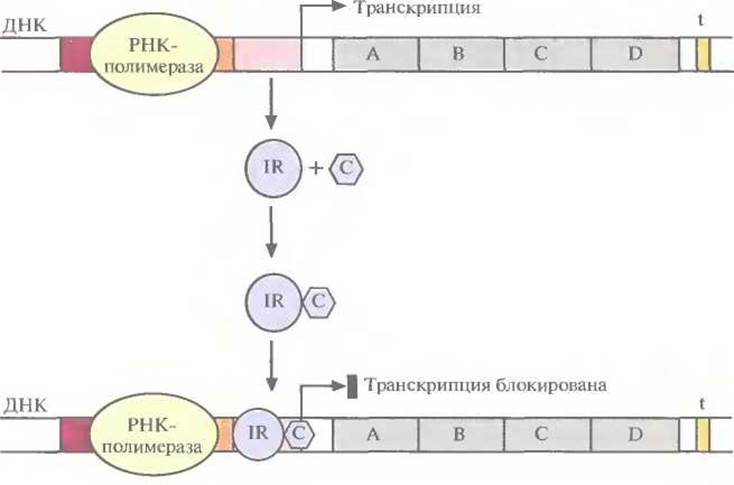
Рис. 3.21. Выключение транскрипции бактериального оперона. Связывание коре прессора (С) с неактивным репрессором (1R) изменяет конформацию последнего. Комплекс корепрессор—репрессор (С—IR) связывается с оператором и блокирует транскрипцию.
Регуляция транскрипции с помощью репрессора называется отрицательной. Если же система регуляции направлена на повышение скорости транскрипции, то она называется положительной. Рассмотрим вкратце этот процесс. Белок-активатор связывается с участком между ТАТА-боксом и сайтом инициации транскрипции. При этом он не только не блокирует перемещение РНК-полимеразы вдоль молекулы ДНК, а напротив, ускоряет его, действуя как своего рода «смазка». Активаторы специфичны для определенных сайтов активации. Иногда с работающим активатором связывается эффектор, переводящий его в неактивную форму; тогда скорость транскрипции уменьшается (рис. 3.22). В других случаях эффектор, напротив, активирует неработающий активатор. Чтобы понять детали регуляции транскрипции у бактерий, необходимо провести тщательный анализ мутаций, которые влияют на данную регуляторную систему, и исследовать in vitro различные сайты связывания белков и ДНК.
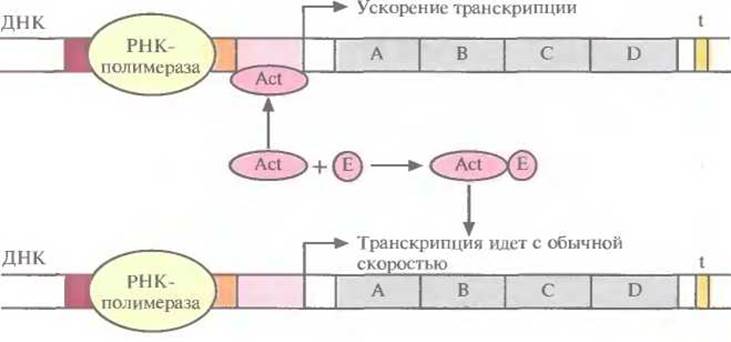
Рис. 3.22. Изменение скорости транскрипции. Активатор (Act) связан с участком между ТАТА-боксом и сайтом инициации транскрипции, скорость транскрипции повышена. Эффектор (Е) связывается с активатором и препятствует его соединению с ДНК; скорость транскрипции уменьшается
