Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Молекулярная биотехнология микробиологических систем
Бактерии, стимулирующие рост растений
Гидрогеназа
Нежелательная побочная реакция фиксации азота — восстановление нитрогеназой Н+ до Н2 (газообразный водород), в ходе которой энергия (в форме АТР) расходуется на образование водорода, который в конечном счете просто улетучивается. В результате только от 40 до 60% всего потока электронов, проходящих через нитрогеназный комплекс, передается на N2, что значительно уменьшает эффективность процесса фиксации азота. В принципе, если бы Н2 мог превратиться обратно в Н+, потери энергии были бы ниже, и процесс фиксации азота стал бы более эффективным. Устранить же эту побочную реакцию прямым путем невозможно, поскольку она обусловлена особенностями химического строения активного центра нитрогеназы, и если попытаться блокировать ее, изменив структуру фермента, то неизбежно произойдет и уменьшение активности нитрогеназы.
Метаболизм водорода
В середине 1970-х годов было показано, что некоторые штаммы Bradyrhizobium japonicum могут расти в микроаэрофильных условиях (при низкой концентрации кислорода), используя в качестве источника энергии водород. Для этого они синтезируют фермент гидрогеназу, способную превращать атмосферный Н2 в Н+ (рис. 14.5). Чтобы проверить, можно ли с помощью этих штаммов влиять на рост сои, растения инфицировали В. japonicum, синтезирующими гидрогеназу (Нuр+). Растения давали большую биомассу и усваивали больше азота, чем те, которые были заражены Hup--штаммами, даже несмотря на более высокий уровень нитрогеназной активности последних (табл. 14.3). По результатам этого и аналогичных экспериментов был сделан вывод, что наличие системы ассимиляции водорода у симбиотических диазотрофов типа В. japonicum повышает их способность стимулировать рост растений, по-видимому, в результате связывания и рециркуляции газообразного водорода, образующегося в клубеньках при участии нитрогеназы (рис. 14.5).
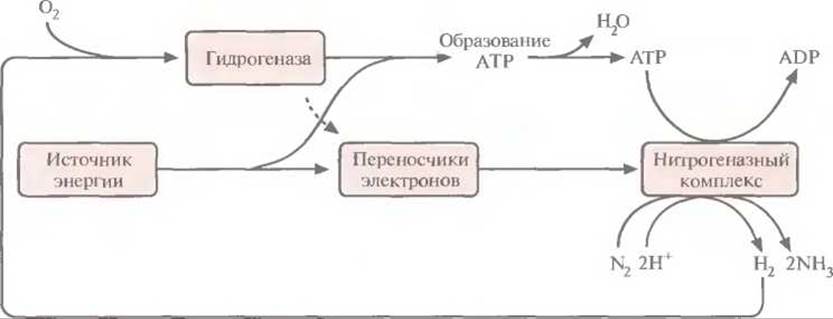
Рис. 14.5. Рециркуляция газообразного водорода — побочного продукта фиксации азота. Нитрогеназа катализирует образование водорода, используя энергию гидролиза АТР, а гидрогеназа катализирует его утилизацию.
Несмотря на выгоды, которые получает растение от симбиоза с диазотрофным микроорганизмом, обладающим системой повторного использования водорода, в природных условиях такая система при участии штаммов Rhizobium встречается редко. Согласно результатам тестирования, представленным в табл. 14.4, большинство рассмотренных природных штаммов Rhizobium и Bradyrhizobium имеют фенотип Hup-. Было проверено по несколько штаммов каждого из указанных видов, а для В. japonicum их было более 1400. Ясно, что как только удастся достаточно подробно изучить генетическую природу гидрогеназной системы и идентифицировать соответствующие гены, коммерческие Hup--штаммы Rhizobium будут первыми кандидатами на превращение в штаммы с фенотипом Нuр+.
Модификация генов гидрогеназ
На изучение гидрогеназ как диазотрофных, так и недиазотрофных микроорганизмов в последниe 20 лет было затрачено много усилий, и тем не менее строение и функции этих ферментов до конца не установлены. Многие микроорганизмы синтезируют более одной гидрогеназы, при этом часто они состоят больше чем из одной полипептидной цепи. Одни гидрогеназы только связывают атмосферный водород, в то время как другие при соответствующих условиях могут также синтезировать его. Из всего этого следует, что вряд ли для преобразования штамма Hup- Rhizobium в Нuр+ будет достаточно простого включения в его геном гена одной из гидрогеназ. Включенный ген(ы) должен кодировать все субъединицы фермента, который должен быть совместим с электронтранспортной системой организма-хозяина.
Таблица 14.3. Относительная активность нитрогеназы и гидрогеназы и способность В. japonicum Hup+ (SR) и трех Нuр- -мутантов (SR1, SR2 и SR3) стимулировать рост растений1) 2)
|
Штамм В. japonicum |
Относительная активность нитрогеназы |
Относительная активность гидрогеназы |
Относительная сухая масса растения |
Относительное содержание азота |
|
SR |
1,00 |
1,00 |
1,00 |
1,00 |
|
SR1 |
1,27 |
0,01 |
0,81 |
0,93 |
|
SR2 |
1,13 |
0,01 |
0,74 |
0,91 |
|
SR3 |
1,23 |
0,01 |
0,65 |
0,85 |
1) Из работы Albrecht et al. Science 203: 1255—1257, 1979.
2) Активность нитрогеназы оценивали по зависимости количества ацетилена, восстановленного до этилена, от времени; активность гидрогеназы определяли при помощи водородного электрода. Сухая масса растения включает массу листьев и корней. Содержание азота рассчитывали как долю сухой массы, приходящуюся на азот. Все величины нормированы относительно таковых для родительского штамма.
Таблица 14.4. Доля природных штаммов Rhizobium и Bradyrhizobium, у которых есть система ассимиляции водорода (Нuр+)1)
|
Бактерия Штаммы Нuр+, % |
|
|
Rhizobium leguminosarum bv. leguminosarum |
9,3 |
|
Rhizobium meliloti |
21 |
|
Rhizobium leguminosarum bv. trifolii |
0 |
|
Rhizobium leguminosarum bv. phaseoli |
0 |
|
Bradyrhizobium japonicum |
21 |
|
Bradyrhizobium sp. |
91 |
1) Из работы Evans et аl. Літи. Rev. Microbiol. 41: 335—361, 1987
Наиболее распространенная стратегия выделения генов гидрогеназ — генетическая комплементация. Первый из таких генов, ген мембраносвязанной гидрогеназы Е. coli, был идентифицирован методом комплементации у мутантной Е. coli, неспособной синтезировать активную гидрогеназу, с использованием банка клонов ДНК Е. coli дикого типа, созданного с помощью плазмиды pBR322. Мутант, содержащий дефектную мембраносвязанную гидрогеназу, не рос на минимальной среде в присутствии формиата, при этом активность эндоплазматической гидрогеназы оставалась неизменной. Трансформированные клетки, способные расти на такой среде, проверяли на присутствие в них активной гидрогеназы. Трансформант, у которого активность гидрогеназы восстановилась до такого же уровня, как у штамма дикого типа, содержал плазмиду, кодирующую белок мол. массой примерно 60 000 Да, что соответствует мол.
массе одной из субъединиц мембраносвязанной гидрогеназы Е. coli. Дальнейшие исследования показали, что в гидрогеназную систему Е. coli входит множество генов.
Затем были идентифицированы гидрогеназные гены (hup) B.japonicum; для этого использовался банк клонов ДНК дикого типа, созданный с помощью космидного вектора pLAFRl с широким кругом хозяев, и мутанты Hup- В. japonicum. Присутствие гидрогеназы, связывающей атмосферный водород, в трансформированных мутантных клетках Hup- определяли по способности активного фермента восстанавливать метиленовый синий в атмосфере водорода. Более детальное исследование показало, что hup-гены B.japonicum образуют по крайней мере два, а возможно, и три оперона, охватывающих примерно 15 т.п.н., причем hup-гены Rhizobium leguminosarum аналогичны таковым В. japonicum как в отношении нуклеотидной последовательности, так и в том, что касается организации генов. Таким образом, идентифицированные hup-гены B.japonicum можо использовать в качестве гибридизационных зондов для поиска гомологичных генов из банка клонов R. leguminosarum.
После идентификации huр-генов R. leguminosarum, несмотря на всю сложность гидрогеназной системы, удалось «переместить» ее из Hup + -штамма R. leguminosarum в штамм Hup- (табл. 14.5). Растения бобов, на которых образовывали клубеньки бактерии рекомбинантного Нuр+-штамма R. leguminosarum, росли быстрее и содержали больше азота, чем растения, инокулированные Нuр--штаммом (табл. 14.5).
Работы по исследованию генов гидрогеназ не вызвали столь большого интереса, как исследования nif-генов, и тем не менее они убедительно продемонстрировали целесообразность применения методов генной инженерии для повышения способности диазотрофных микроорганизмов стимулировать рост растений. Теперь нужно проверить, приведет ли введение hup-генов в геномы других диазотрофных микроорганизмов (как несимбиотических, так и симбиотических) к такому же эффекту.
Гидрогеназная система может применяться не только для повышения эффективности фиксации азота. Так, очищенную гидрогеназу можно использовать для преобразования и запасания солнечной энергии; регенерации кофакторов, принимающих участие в промышленных ферментативных процессах; синтеза специфических химических соединений, требующего участия Н2 в качестве восстановителя; для удаления трития из воды, которая использовалась для охлаждения реакторов атомных электростанций; для синтеза Н2 из органических отходов; получения водородно-кислородных топливных ячеек. Однако, несмотря на то что уже идентифицировано и охарактеризовано более дюжины генов гидрогеназ, пока ни один из них не использовался для крупномасштабного синтеза этих ферментов.
Таблица 14.5. Рост растений и ассимиляция азота после введения генов hup в Hup- -штамм R. legumirmsarum1)
|
Фенотип |
Относительная сухая масса растения |
Относительное содержание азота |
Относительная площадь листа |
Относительная концентрация азота |
|
Hup- |
1,00 |
1,00 |
1,00 |
1,00 |
|
Hup+ |
1,35 |
1,52 |
1,53 |
1,15 |
1) Из работы Brewin, Johnston, U.S. patent 4,567,146, январь 1986.