Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Молекулярная организация клеток
Плазматическая мембрана
Мембранные углеводы
На поверхности всех эукариотических клеток имеются углеводы. Они представлены в виде олигосахаридных и полисахаридных цепей, ковалентно присоединенных к мембранным белкам (гликопротеины) и к липидам (гликолипиды). Масса углеводов плазматической мембраны колеблется от 2 до 10% от массы мембраны. Большинство белков плазматической мембраны, выступающих на поверхности клеток, связаны с остатками Сахаров. В то же время из десяти липидных молекул в наружном монослое большинства плазматических мембран с углеводами связана менее чем одна молекула (см. разд. 6.1.6). Пятидесятикратное превышение в мембране числа липидных молекул над молекулами белка означает, что липидных молекул, связанных с углеводами в обычной (типичной) мембране больше, чем белковых. Однако такой гликопротеин как гликофорин может иметь большое количество боковых олигосахаридных цепей, а каждая молекула гликолипида - лишь одну. Кроме того, многие плазматические мембраны содержат молекулы интегральных протеогликанов. Протеогликаны состоят из длинных полисахаридных цепей, присоединенных к белковому кору, и выявляются главным образом на внешней стороне клетки как часть внеклеточного матрикса. Однако в некоторых случаях кор интегральных протеогликанов, по-видимому, пронизывает липидный бислой.
6-18
6.3.1. Углеводы в биологических мембранах располагаются только на поверхности, не контактирующей с цитозолем [18]
Как мы уже знаем, биологические мембраны чрезвычайно асимметричны: наружный и внутренний монослои различаются как по липидному составу, так и по белковому. Такая же асимметрия наблюдается и в распределении углеводов; углеводные цепи основной массы гликолипидов, гликопротеинов и протеогликанов во внутренних и плазматических мембранах локализованы исключительно на той стороне мембраны, которая не контактирует с цитозолем. В плазматических мембранах остатки Сахаров выступают на внешнюю поверхность клетки, а во внутренних мембранах они обращены внутрь ограниченного мембраной компартмента. Существуют два различных варианта присоединения олигосахаридов к мембранным гликопротеинам: они могут быть «пришиты» N-связью к остаткам аспарагина в полипептидной цепи или О-связью к остаткам серина или треонина. N-связанные олигосахариды обычно содержат около 12 Сахаров и строятся на основе общего ядра, состоящего из остатков маннозы. О-связанные олигосахариды, как правило, короче (длиной около 4 сахарных остатков).
Одним из простейших способов демонстрации присутствия Сахаров на клеточной поверхности является использование белков, связывающих углеводы, и названных лектинами. Существует ряд белков, обладающих сайтами, узнающими и связывающими специфические последовательности сахарных остатков. Первоначально они были выделены из семян растений. Некоторые лектины чрезвычайно токсичны и служат для отпугивания животных, которые могли бы съесть семена. Совсем недавно было показано, что лектины имеются не только у растений, но и у многих других существ, включая животных. Некоторые из них находятся на поверхности клеток, и, по-видимому, участвуют в межклеточном узнавании (см. рис. 5-42). Поскольку лектины связываются с гликопротеинами, протеогликанами и гликолипидами, находящимися на поверхности клеток, они широко используются в клеточной биологии в качестве биохимического маркера для локализации и выделения молекул плазматической мембраны, содержащих сахара. В таблице 6-2 представлены наиболее часто используемые растительные лектины и указана их специфичность по отношению к сахарам.
Таблица 6-2. Наиболее популярные коммерческие препараты растительных лектинов и сахара, которые ими узнаются
|
Лектины |
Сахароспецифичность |
|
Конканавалин А |
a-D-глюкоза и a-D-манноза |
|
Лектин из сои |
а-галактоза и N-ацетил-D-галактозамин |
|
Лектин из зародышей пшеницы |
N-ацетилглюкозамин |
|
Лектин из семян лотоса |
Фукоза |
Термин клеточная оболочка, или гликокаликс, часто используются для обозначения обогащенной углеводами периферической зоны на поверхности большинства эукариотических клеток. При использовании меченых лектинов или различных красителей, например рутениевого красного, эта зона отчетливо видна на электронных микрофотографиях (рис. 6-39). Хотя углеводы главным образом присоединены к молекулам, входящим в состав плазматической мембраны, гликокаликс может также содержать гликопротеины и протеогликаны, которые секретируются клетками и затем адсорбируются на клеточной поверхности (рис. 6-40). Некоторые из этих адсорбированных макромолекул являются компонентами внеклеточного матрикса. Так что вопрос о том, где кончается плазматическая мембрана и начинается внеклеточный матрикс, можно считать чисто семантическим.
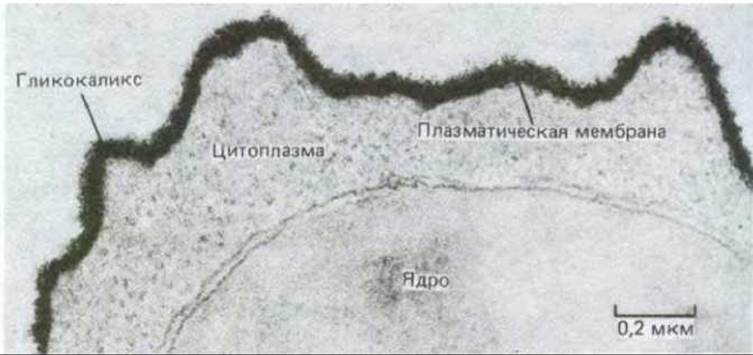
Рис. 6-39. Электронная микрофотография поверхности лимфоцита, окрашенного рутениевым красным с целью получить контрастное изображение клеточной оболочки (гликокаликса). (С любезного разрешения А. М. Glauer и G. М. W. Cook.)
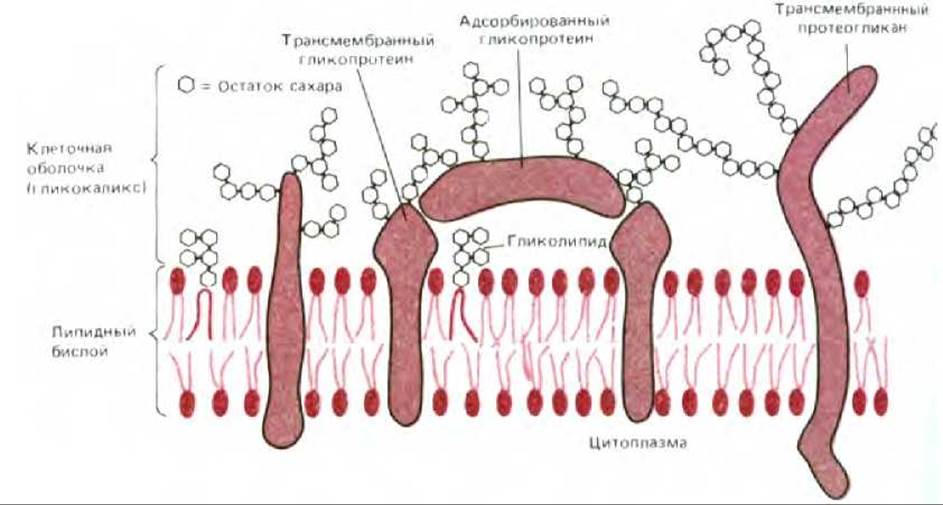
Рис. 6-40. Схематическое изображение клеточной оболочки (гликокаликса), состоящей из боковых олигосахаридных цепей гликолипидов и интегральных мембранных гликопротеинов и полисахаридных цепей протеогликанов. В некоторых клетках присутствуют также адсорбированные гликопротеины и протеогликаны (не показаны). Обратите внимание, что все углеводы располагаются на наружной стороне мембраны. Некоторые интегральные гликопротеины и протеогликаны могут быть ковалентно связаны через специфические олигосахариды с фосфатидилинозитолом, находящимся во внешнем монослое плазматической мембраны (см. рис. 6-14.)
Ясно, что высокая концентрация углеводов на клеточной поверхности должна оказывать существенное влияние на многие функции плазматической мембраны. Однако природа этого влияния еще не понята. Структурная сложность некоторых олигосахаридов, а также то, что углеводы располагаются только на поверхности клетки, свидетельствуют о важной роли углеводов в процессах межклеточного узнавания и узнавания между клеткой и матриксом. Для некоторых случаев существуют веские доказательства этой роли (см. разд. 15.4.2 и 15.4.9), однако гораздо чаще такие функции углеводов клеточной поверхности однозначно доказать очень трудно.
Заключение
В плазматических мембранах всех эукариотических клеток большинство белков, расположенных на поверхности клетки, а также некоторые липидные молекулы наружного липидного монослоя ковалентно связаны с олигосахаридными цепями. Некоторые плазматические мембраны содержат молекулы интегральных протеогликанов, в которых несколько полисахаридных цепей ковалентно сшиты с трансмембранным белком или связанным с липидами коровым белком. Хотя функция углеводов клеточной поверхности пока непонятна, представляется вероятным, что по крайней мере некоторые из них принимают участие в процессах межклеточного узнавания и узнавания между клеткой и матриксом.