Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Нервная система
Клетки нервной системы: строение и функция
Нервная система обеспечивает быструю связь между отдаленными частями тела. Благодаря своей роли коммуникативной сети нервная система управляет реакциями на внешние раздражители, перерабатывает информацию и генерирует сложные комплексы сигналов для регулирования сложного поведения. Кроме того, нервная система способна самообучаться: по мере обработки и запоминания сенсорной информации о внешнем мире происходит надлежащая «подстройка» нервной системы, в результате чего изменяется характер последующих действий.
Основные пути передачи нервных сигналов были прослежены более ста лет назад, еще до того как стала понятна роль отдельных нервных клеток. На рис. 19-1 показан общий план организации нервных связей. Подобно большому компьютеру, нервная система позвоночного состоит из главного процессора - центральной нервной системы, включающей головной и спинной мозг, - который с помощью «проводов», т. е. нервов, соединен с многочисленными периферическими структурами: органами чувств, доставляющими входную информацию, и мышцами (а также в меньшей степени железами), реализующими выходные команды. Имеются также связи с группами периферических нервных клеток-ганглиями, которые в одних случаях просто поддерживают коммуникацию между периферией и центром, а в других служат вспомогательными мини-компьютерами. У беспозвоночных имеется сходная организация, но центральная нервная система у них развита слабее, тогда как ганглии играют более значительную роль и более автономны.
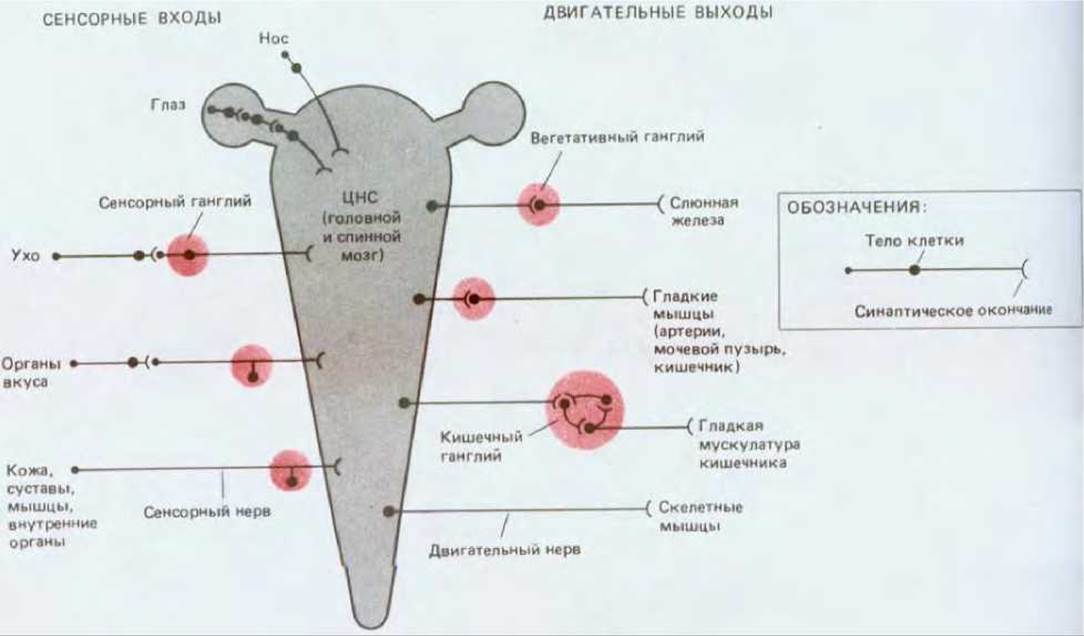
Рис. 19-1. Сильно упрощенная схема организации нервной системы позвоночного. Показано, как сенсорная информация передается с периферии в центральную нервную систему (ЦНС), а двигательные команды - из ЦНС на периферию. Передачу сенсорных и двигательных сигналов осуществляют нервные клетки, тела которых (показаны большими черными точками) во многих случаях сгруппированы в ганглиях (цветные кружки) за пределами ЦНС, а аксоны объединены в пучки и образуют нервы (черные линии). Нервы, ганглии и органы чувств вместе составляют периферическую нервную систему. Некоторые ганглии служат просто «передаточными станциями», а другие (особенно это касается вегетативных кишечных ганглиев, регулирующих перистальтику) представляют собой сложные системы связанных между собой нейронов, способных функционировать даже без всякого участия ЦНС. Вставочные нейроны, входящие в состав центральной нервной системы, не показаны.
У разных животных детальная структура нервных связей чрезвычайно сильно варьирует в пределах общей схемы; однако свойства отдельных нейронов во многом сходны независимо от того, идет ли речь о моллюсках, насекомых, амфибиях или млекопитающих.
19.1.1. Функция нервной клетки определяется длиной ее отростков [2]
Фундаментальная задача, стоящая перед нейроном, состоит в приеме и передаче сигналов. Выполнять эти функции нейрону позволяют его необычные размеры и форма: длина нервной клетки человека, соединяющей, например, спинной мозг с какой-либо мышцей стопы, может достигать одного метра. Каждый нейрон состоит из тела клетки (содержащего ядро) и расходящимися от него длинными тонкими отростками. Обычно это один длинный аксон, проводящий сигналы от тела клетки к отдаленным мишеням, и несколько более коротких ветвящихся дендритов, которые, подобно антеннам, принимают сигналы от аксонов других нервных клеток (рис. 19-2). Тело нейрона тоже принимает сигналы. Отдаленный конец аксона обычно ветвится, что позволяет передавать сигнал одновременно нескольким клеткам-мишеням. Степень ветвления дендритов тоже может быть очень высокой - в некоторых случаях один нейрон способен принимать до 100000 сигналов. Характер ветвления аксонов и дендритов у нейронов функционально различных типов может быть поразительно разнообразным (рис. 19-3).
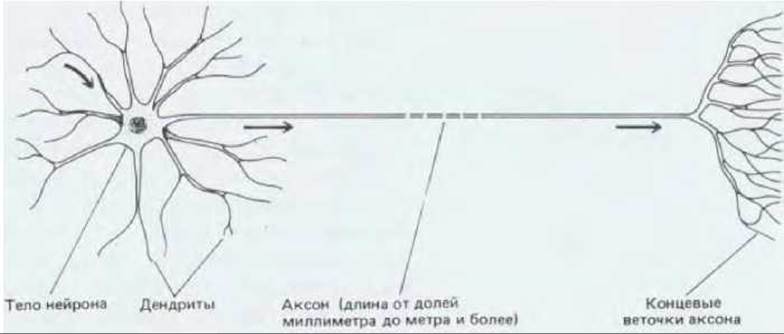
Рис. 19-2. Схематическое изображение типичного нейрона позвоночных. Стрелками указано направление передачи сигналов. Самые крупные нейроны достигают у человека длины около метра и имеют аксоны диаметром до 15 мкм.
19.1.2. Нервные клетки передают электрические сигналы [3]
Значение сигналов, передаваемых нервной клеткой, зависит от того, какую роль играет эта клетка в работе нервной системы в целом. В моторных (двигательных) нейронах сигналы служат командами для сокращения определенных мышц. В сенсорных (чувствительных) нейронах они передают информацию о раздражителях определенного та таких как свет, механическая сила или химическое вещество, воздействующих на тот или иной участок тела. В интернейронах (вставочных нейронах), связывающих один нейрон с другим, сигналы обеспечивают сложное взаимодействие и объединение информации из нескольких различных источников и участвуют в регуляции сложного поведения.
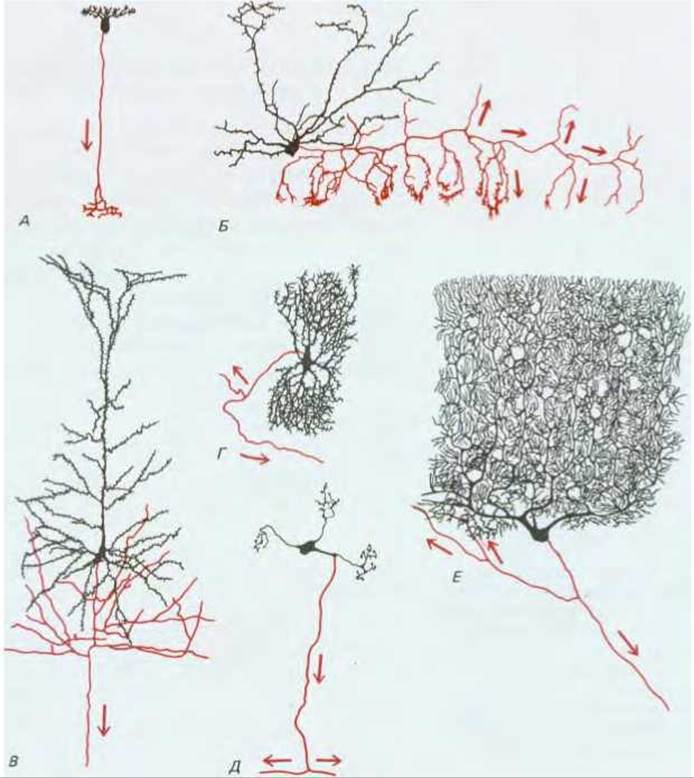
Рис. 19-3. Некоторые из многочисленных типов нервных клеток позвоночных, как они выглядят после окраски по Гольджи. Эта методика, включающая погружение ткани в раствор солей металлов, позволяет полностью окрашивать в черный цвет небольшую долю клеток, имеющихся в препарате, и дает возможность увидеть все разветвления клеточных отростков. От тела нейрона отходит множество дендритов, получающих входные сигналы от других клеток, и один тонкий ветвящийся аксон, передающий выходные сигналы в направлении, показанном стрелками. Аксоны представлены красным цветом, тело клетки и дендриты - черным. У клеток А и Б короткие аксоны, они изображены здесь полностью. У клеток В - Е аксоны очень длинные, и на рисунке показаны только их начальные участки. А - биполярная клетка из сетчатки ящерицы; Б- корзинчатая клетка из мозжечка мыши; В-пирамидная клетка из коры головного мозга кролика; Г-нейрон из ствола мозга человека; Д-одна из клеток-зерен мозжечка кошки; Е-клетка Пуркинье из мозжечка человека. Эта последняя клетка, имеющая широко разветвленную сеть дендритов, получает сигналы от более чем 100000 других нейронов; она представляет собой элемент мозгового механизма, регулирующего сложные движения. Рисунки сделаны в разных масштабах: длина биполярной клетки (А) около 100 мкм, тогда как изображенная на рисунке часть клетки Пуркинье (Е) имеет ширину около 400 мкм (длина ее аксона достигает нескольких сантиметров).
Несмотря на различное значение сигналов, природа их во всех случаях одинакова и состоит в изменении электрического потенциала плазматической мембране нейрона. Связь осуществляется благодаря тому, что электрическое возмущение, возникшее в одном участке клетки, распространяется на другие участки. Эти возмущения затухают по мере удаления от их источника, если нет дополнительного усиления на пути следования сигнала. На коротких расстояниях затухание незначительно, и многие небольшие нейроны проводят сигналы пассивно, без усиления. Однако для дальней связи такого пассивного распространения недостаточно. Поэтому нейроны с более длинными отростками используй активный сигнальный механизм, составляющий одно из самых удивительных и характерных свойств нейрона. Электрический стимул, сила которого превышает определенную пороговую величину, вызывает «вспышку» электрической активности, распространяющейся с большой скоростью вдоль плазматической мембраны нейрона и поддерживаемо с помощью автоматического усиления на протяжении всего пути. Эта бегущая волна электрического возбуждения, называемая потенций» действия или нервным импульсом, способна передавать информацию без затухания от одного конца нейрона к другому со скоростью до 100 м/с и более.
19.1.3. Связь между нейронами осуществляется в синапсах с помощью химических сигналов [4] |
Сигналы, проводимые нейронами, передаются от одной клетки к другой в особых местах контакта, называемых синапсами. Обычно передача осуществляется, как это ни странно на первый взгляд, непрямым путем. Клетки электрически изолированы друг от друга: пресинаптическая клетка отделена от постсинаптической промежутком - синаптической щелью. Изменение электрического потенциала в пресинаптической клетке приводит к высвобождению вещества, называемого нейромедиатором, которое хранится в ограниченных мембраной синаптических пузырьках и высвобождается путем экзоцитоза. Нейромедиатор диффундирует через синаптическую щель и вызывает изменение электрофизиологического состояния постсинаптической клетки (рис. 19-4). Как мы увидим позже, механизм передачи сигнала через такие химические синапсы более гибок и доступен для адаптации, чем прямая электрическая связь, осуществляемая через щелевые контакты (разд. 14.1.7), которая тоже используется, но гораздо реже.
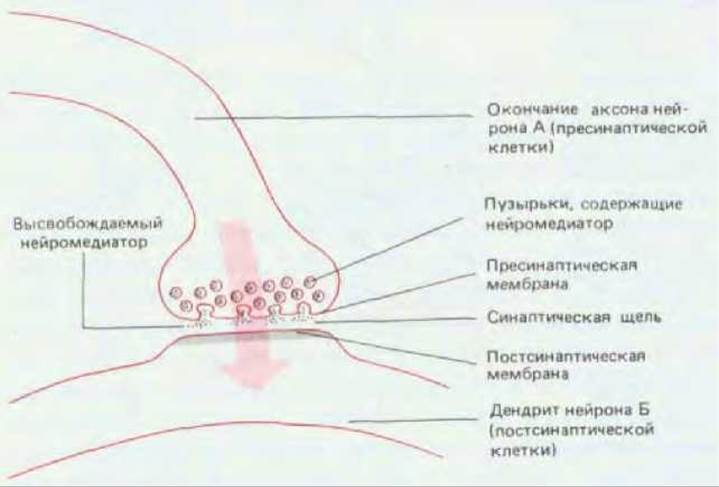
Рис. 19-4. Схема типичного синапса. Электрический сигнал, приходящий в окончание аксона клетки А, приводит к высвобождению в синаптическую щель химического посредника (нейромедиатора), который вызывает электрическое изменение в мембране дендрита клетки В. Широкая стрелка указывает направление передачи сигнала.
Химический синапс - это место интенсивной биохимической активности, включающей распад, обновление и секрецию белков и других молекул. Однако биохимическим центром нейрона служит тело клетки, где заложены основные «инструкции» по синтезу белка. Поэтому нейрону необходима эффективная внутриклеточная система транспорта молекул из тела клетки к самым отдаленным участкам аксона и дендритов. Как же организована эта транспортная система и какие молекулы переносятся в действительности?
19.1.4. Вновь синтезируемые материалы переносятся из тела нервной клетки в аксоны и дендриты с помощью механизмов медленного и быстрого транспорта [5]
С помощью электронной микроскопии установлено, что тело типичного крупного нейрона содержит огромное количество рибосом, часть которых находится в цитозоле, а часть прикреплена к мембранам гранулярного эндоплазматического ретикулума (рис. 19-5, А). Хотя дендриты часто содержат небольшое количество рибосом, в аксоне их нет, и поэтому белки аксона должны синтезироваться на рибосомах тела клетки (рис. 19-5, Б). Потребности аксона значительны: например, толщина большого мотонейрона человека может достигать 15 мкм при длине 1 м, что соответствует объему примерно 0,2 мм3, а это почти в 10000 раз больше объема печеночной клетки. Поскольку в таком нейроне только одно ядро, соотношение цитоплазмы к ДНК в нейроне намного выше, чем в любой другой клетке человеческого организма.
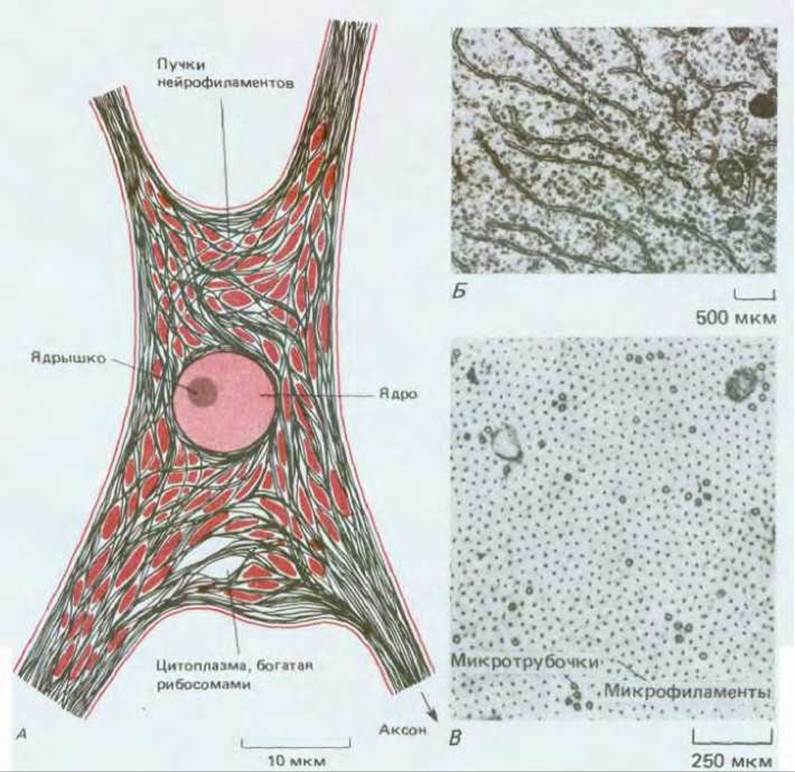
Рис. 19-5. Строение цитоплазмы типичного крупного нейрона (мотонейрона из спинного мозга). А. Схематическое изображение тела клетки при небольшом увеличении; видно, что участки цитоплазмы, богатые рибосомами, расположены между пучками нейрофиламентов и других белков цитоскелета. Б. Электронная микрофотография одного из таких богатых рибосомами участков; которые рибосомы прикреплены к гранулярному эндоплазматическому ретикулуму, другие ни с чем к связаны. В. Электронная микрофотография части поперечного среза аксона; можно видеть большое число микрофиламентов и микротрубочек, но рибосомы отсутсвуют. Мембранные пузырьки передвигаются, вероятно, вдоль ближайших микротрубочек с помощью механизма быстрого аксонного транспорта. (С любезного разрешения Jennifer La Vail (Б) и John Hopkins (В).)
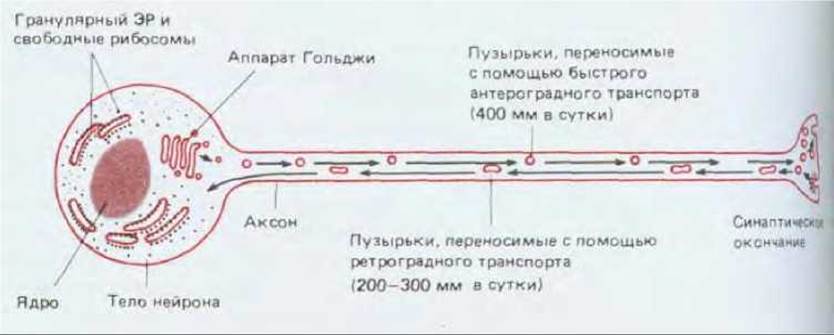
Рис. 19-6. Нейрон как секреторная клетка, у которой место секреции (окончание аксона) расположено на большом расстоянии от места синтеза макромолекул (тела клетки). При такой организации необходим механизм быстрого аксонного транспорта. Из приведенной схемы не следует заключать, что все синаптические пузырьки транспортируются из тела нейрона: в большинстве нейронов они образуются в основном путем повторного использования мембраны в окончании аксона.
В аксоне в наибольших количествах содержатся белки, образующие микротрубочки, нейрофиламенты (класс промежуточных филаментов) и актиновые филаменты (рис. 19-5, В). Белки цитоскелета доставляются из тела клетки и движутся по аксону со скоростью от 1 до 5 мм в сутки. Это медленный аксонный транспорт (подобный вид транспорта имеется и в дендритах, содержащих несколько иной набор белков, связанных с микротрубочками - см. разд. 11.4.7). Другие белки цитозоля, включая многие ферменты, тоже переносятся с помощью медленного аксонного транспорта, механизм которого не ясен.
Нецитозольные материалы, необходимые в синапсе, такие как секретируемые белки и мембраносвязанные молекулы, перемещаются от тела клетки с помощью гораздо более быстрой разновидности аксонного транспорта. Эти белки и липиды переносятся от мест их синтеза в эндоплазматическом ретикулуме к аппарату Гольджи, расположенному вблизи ядра (часто у основания аксона). Отсюда эти молекулы, «упакованные» в мембранные пузырьки, переносятся путем быстрого аксонного транспорта со скоростью до 400 мм в сутки вдоль путей, образуемых в аксоне и дендритах микротрубочками (разд. 11.4.8). Таким же образом транспортируются митохондрии. Так как этим способом в аксонах и в дендритах перемещаются разные виды белков, полагают, что транспортируемые молекулы распределяются в теле клетки по различным транспортным пузырькам определенных типов (разд. 8.9.4).
К белкам, быстро переносимым по аксону, относятся и белки, предназначенные для высвобождения в синапсе, такие как нейропептиды, выделяемые многими нейронами в качестве нейромедиаторов, часто в сочетании с небелковыми медиаторами. С точки зрения внутренней организации нейроны можно представить как секреторные клетки, в которых место выделения секрета находится на громадном расстоянии от места образования белков и мембран (рис. 19-6).
19.1.5. Благодаря ретроградному транспорту поддерживается обратная химическая связь между окончаниями и телом нервной клетки [5, 6]
Быстрый аксонный транспорт необходим во время развития клетки для роста аксонов и дендритов, которые удлиняются путем добавления новой мембраны на их концах. Быстрый аксонный транспорт имеется и в нейроне, закончившем рост, у которого количество мембранного материала в кончиках отростков не увеличивается. В этом случае быстрый транспорт мембран от тела клетки, называемый антероградным, должен быть точно сбалансирован с быстрым ретроградным транспортом мембран в обратном направлении - от концов клеточных отростков. Механизмы двух встречных направлений быстрого транспорта сходны, но не идентичны. Быстрый ретроградный транспорт, скорость которого в два раза меньше скорости быстрого антероградного транспорта, осуществляется с помощью иных двигательных белков (разд. 10.4.9) и используется для переноса пузырьков несколько большей величины. Структуры, возвращающиеся в тело клетки, состоят частично из стареющих цитоплазматических органелл, например митохондрий, а частично из пузырьков, образующихся при интенсивном эндоцитозе, необходимом для восстановления мембраны в окончании аксона после высвобождения нейромедиатора (см. рис. 19-20). Молекулы, находящиеся во внеклеточной среде вокруг окончания аксона, могут захватываться этими эндоцитируемыми пузырьками и вместе с ними переноситься к телу клетки. Таким образом, биосинтетический аппарат, находящийся в теле клетки, способен «узнавать» об изменениях обстановки у окончания аксона и, как мы увидим позже, соответственно реагировать на них (разд. 19.7.10).
Ретроградный транспорт позволяет тем, кто изучает анатомию нервной системы, легко прослеживать нервные связи с помощью несложной методики, показанной на рис. 19-7.
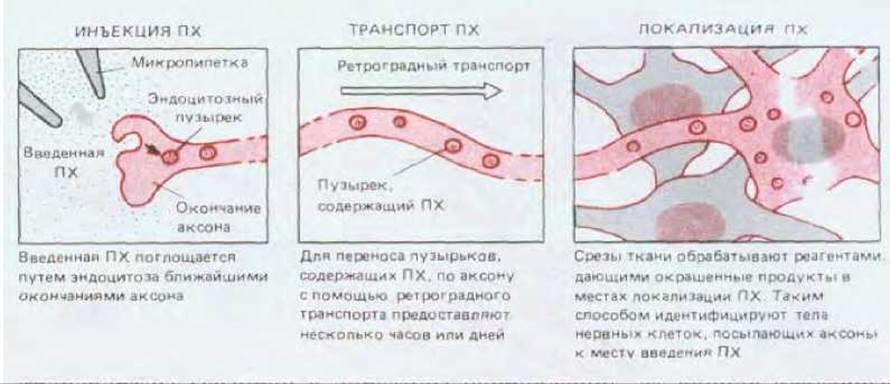
Рис. 19-7. Использование быстрого аксонного транспорта для идентификации и определения локализации отдаленных нервных клеток, аксоны которых оканчиваются в исследуемом участке. В качестве маркера наиболее широко используется фермент пероксидаза хрена (ПХ), так как его молекулы могут быть обнаружены в очень малых количествах по окрашенным продуктам реакции, катализируемой этим ферментом.
19.1.6. Нейроны окружены глиальными клетками различного типа [7]
Вся нервная ткань, как периферическая, так и центральная, состоит из клеток двух основных классов. Главная роль принадлежит нейронам, но глиальные клетки, поддерживающие нейроны, превосходят их по численности: в мозгу млекопитающих их примерно в 10 раз больше, чем нейронов. Глиальные клетки окружают нейроны (как их тела, так и отростки) и заполняют пространство между ними. Наиболее изучены шванновские клетки из периферических нервов позвоночных и олигодендроциты из центральной нервной системы позвоночных. Эти клетки обвиваются вокруг аксонов, образуя изоляционный слой в виде миелиновой оболочки (разд. 19.2.4). Три других типа глиальных клеток цетральной нервной системы - это микроглия, эпендимные клетки и астроциты (рис. 19-8). Микроглия относится к несколько обособленному классу: эти клетки функционально близки к макрофагам (разд. 17.5.1) и, подобно им, происходят из кроветворной ткани. Все остальные глиальные клетки имеют общее эмбриональное происхождение с теми нейронами, с которыми они связаны, однако в отличие от большинства нейронов глия, как правило, не способна к электрическому возбуждению. Кроме того, в то время как нейроны после дифференцировки уже не могут делиться, большая часть глиальных клеток сохраняет эту способность на протяжении всей жизни.
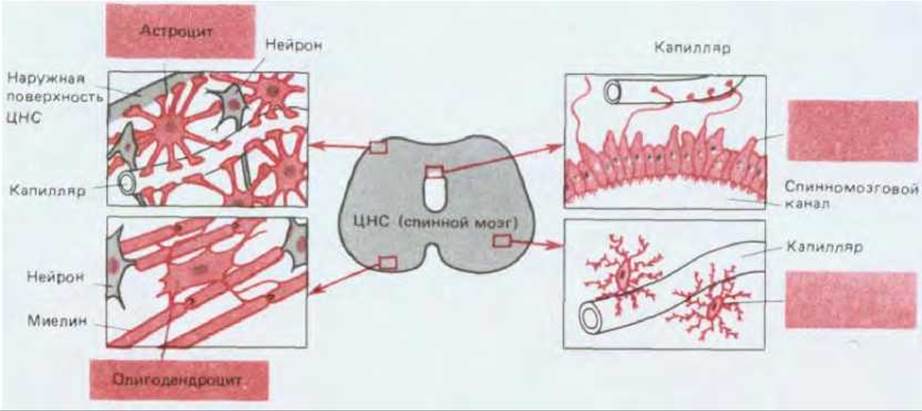
Рис. 19-8. Три основных класса глиальных клеток из центральной нервной системы позвоночных. Глиальные клетки выделены цветом. Астроциты, которые наиболее многочисленны, имеют множество радиально отходящих отростков. Некоторые из этих отростков оканчиваются на поверхности нейронов, а другие, с расширенными концами, образуют наружный поверхностный слой ЦНС, который окружает ее кровеносные сосуды и совместно с эндотелиальными клетками капилляров создает гематоэнцефалический барьер. Эпендимные клетки образуют ресничный эпителий, выстилающий центральные полости ЦНС, и отростки этих клеток, так же как и отростки астроцитов, часто оканчиваются на кровеносных сосудах. Олигодендроциты образуют вокруг аксонов ЦНС изолирующую миелиновую оболочку. Микроглиальные клетки по своим функциям и происхождению близки к макрофагам; они участвуют в реакции ткани на повреждение и инфекцию. Эти клетки обычно находятся вблизи кровеносных сосудов.
Эпендимные клетки выстилают внутренние полости головного и спинного мозга (рис. 19-8), а их эпителиальная организация напоминает нам о происхождении центральной нервной системы из эпителиальной трубки (разд. 19.7.1).
Астроциты (рис. 19-8) - самые многочисленные и разнообразные глиальные клетки, но и самые загадочные: их функция все еще в значительной части не выяснена, хотя кажется несомненным, что они играют важную роль в процессе построения нервной системы (разд. 19.7.2) и регулируют химический и ионный состав среды, окружающей нейроны, Например, одна из разновидностей астроцитов имеет отростки с расширенными концами, которые, будучи связаны соединительными комплексами вроде встречающихся в эпителиях (разд. 14.1), образуют изолирующий барьер на внешней поверхности центральной нервной системы. Другие отростки этих же астроцитов образуют сходные «концевые ножки» на кровеносных сосудах, эндотелиальные клетки которых случае капилляров и венул) соединяются здесь необычайно развитыми плотными контактами, так что создается гематоэнцефалический барьер. Этот барьер предотвращает проникновение из крови в ткань мозга водорастворимых молекул, если их не переносят специальные транс портные белки, находящиеся в плазматической мембране эндотелиальных клеток. Таким образом, нейроны оказываются в контролируемой и защищенной среде, что имеет решающее значение для молекулярного механизма передачи электрических сигналов.
Заключение
Нервные клетки, или нейроны, - это клетки с необычайно длинными отростками, передающими электрические сигналы в виде потенциалов действия - бегущих волн электрического возбуждения. Обычно от тела нервной клетки отходит несколько разветвленных дендритов и один длинный аксон. Как правило, сигналы воспринимаются дендритами и телом клетки, а затем распространяются по аксону и передаются другим клеткам в химических синапсах. Здесь электрический сигнал, приходящий в пресинаптическое окончание аксона, индуцирует секрецию нейромедиатора, который в свою очередь вызывает электрическое изменение в постсинаптической клетке.
Нейрон можно рассматривать как секреторную клетку, выделяющую свой секрет - нейромедиатор - на очень большом расстоянии от тем клетки, где синтезируются макромолекулы. Вновь синтезируемые секреторные белки и материал для построения мембраны переносятся по аксону и дендритам благодаря быстрому аксонному транспорту, при котором мелкие мембранные пузырьки движутся вдоль путей, образуемых микротрубочками. Микротрубочки и другие компоненты цитоплазмы, не связанные с мембранами, перемещаются от тела клетки при помощи совершенно другого механизма медленного аксонного транспорта. Быстрый аксонный транспорт осуществляется также и в обратном, ретроградном, направлении, перенося мембранные пузырьки от окончаний аксона к телу клетки.
Нейроны окружены глиальными клетками, которые помогают различным образом регулировать химические и электрические свойства среды, окружающей нейроны.