Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Нервная система
Прием сенсорной информации
Выше было показано, как нервные клетки проводят, перерабатывают и регистрируют электрические сигналы, а затем посылают их мышцам, чтобы вызвать их Сокращение. Но откуда берутся эти сигналы? Имеются два типа источников: спонтанное возбуждение и сенсорные стимулы. Существуют спонтанно активные нейроны, например нейроны мозга, задающие ритм дыхания; весьма сложная картина самопроизвольной активности может генерироваться в одиночной клетке с помощью надлежащих комбинаций ионных каналов тех типов, с которыми мы уже встречались при обсуждении механизмов переработки информации нейронами. Прием сенсорной информации тоже основан на уже известных нам принципах, но в нем участвуют клетки весьма разнообразных и удивительных типов.
Органы чувств должны удовлетворять очень строгим требованиям - с высокой точностью различать стимулы разных типов, воспринимать изменения силы стимула в феноменально широких границах и обладать такой высокой чувствительностью, какую только допускают законы физики. Обонятельная клетка самца непарного шелкопряда способна обнаружить в воздухе одну-единственную молекулу специфического полового аттрактанта (так называемого феромона), выделяемого самкой на расстоянии мили от места нахождения самца. Глаз человека видит и при ярком солнечном свете, и в звездную ночь, когда освещение в 1012 раз слабее, и пять фотонов, поглощенных сетчаткой, воспринимаются как вспышка света.
Мы сконцентрируем внимание на двух органах чувств, в которых клеточные механизмы получения сенсорной информации начинают становиться понятными: на органах слуха и зрения позвоночных. Каждый из этих двух входов в нервную систему содержит высокоспециализированные сенсорные клетки, очень различные, но в обоих случаях обладающие необычайной избирательностью и чувствительностью в широком диапазоне стимулов. Но прежде чем переходить к деталям, полезно будет рассмотреть некоторые общие принципы.
19.6.1. Силу стимула отражает величина рецепторного потенциала [38, 39]
Любой сигнал, получаемый нервной системой, должен прежде всего превратиться в электрический. Превращение сигнала одного вида в другой называется преобразованием, поэтому все сенсорные клетки-преобразователя. В более общем смысле почти каждый нейрон является преобразователем: получая в синапсах химические сигналы, он преобразует их в электрические. Хотя одни сенсорные клетки реагируют на свет, другие на температуру, третьи на определенные химические вещества, четвертые на механическую силу или перемещение и т.д., во всех этих клетках преобразование основано на ряде основных принципов, которые уже рассматривались при обсуждении синаптической передачи с помощью нейромедиаторов. В некоторых органах чувств преобразователь составляет часть сенсорного нейрона, проводящего импульсы, а в других это часть сенсорной клетки, специально приспособленной для преобразования сигнала, но не участвующей в осуществлении дальней связи: такая клетка передает свои сигналы связанному с нею нейрону через синапс (рис. 19-44). Но в обоих случаях воздействие внешнего стимула вызывает в клетке- преобразователе электрический сдвиг, называемый рецепторным потенциалом, который аналогичен постсинаптическому потенциалу и тоже в конечном счете служит для регуляции высвобождения нейромедиатора из другой части клетки.
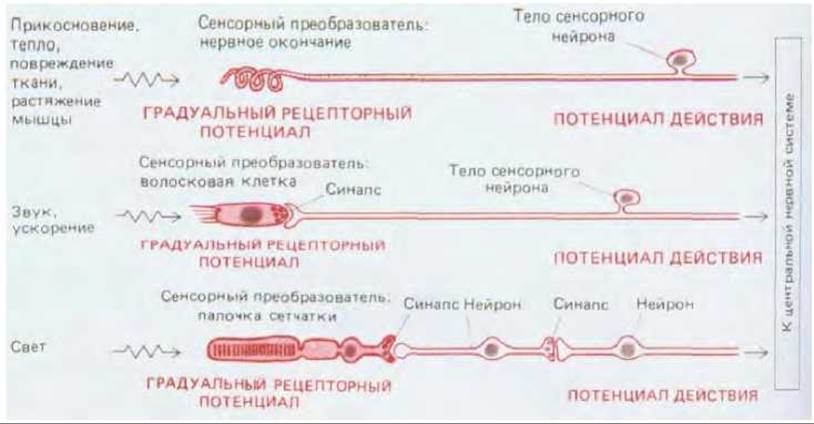
Рис. 19-44. Различные способы передачи сенсорной стимуляции нервной системе. В некоторых случаях сенсорный преобразователь составляет часть нервной системы (верхняя схема); в других случаях это отдельные сенсорные клетки (две нижние схемы). Во всех трех случаях возникающий в Сенсорном преобразователе градуальный рецепторный потенциал преобразуется в частоту импульсов, которые быстро передают сигнал в ЦНС.
Так же как в синапсе, внешние раздражители способны влиять на электрическое состояние клетки как непосредственно, воздействуя на ионные каналы, так и косвенно - через молекулы-рецепторы, запускающие синтез внутриклеточного посредника, который уже воздействует на ионные каналы. Как полагают (хотя здесь еще не все ясно), слуховые сенсорные клетки используют прямой механизм действия с участием рецепторов, связанных с каналами, а сенсорные клетки глаза - непрямой путь через рецепторы, связанные с G-белком.
19.6.2. Волосковые клетки внутреннего уха реагируют на отклонение стереоцилий [40]
Ухо предназначено не только для слуха: внутреннее ухо воспринимает также информацию о направлении силы тяжести и об ускоренном движении, поэтому оно необходимо для сохранения равновесия и координации движений. В основе всех сенсорных функций уха лежит механорецепция, а именно улавливание небольших смещений среды, окружающей сенсорные клетки. В случае звуковых волн эти смещения представляют собой быстрые вибрации, а восприятие ускоренного движения или силы тяжести связано с более медленными и плавными перемещениями. Все клетки внутреннего уха, ответственные за различные виды механорецепции, имеют характерное строение: на верхней поверхности такой клетки находится пучок гигантских микроворсинок, получивших название стереоцилий (рис. 19-45; см. также разд. 11.6.10). Поэтому такие клетки называют волосковыми.
У высших позвоночных все волосковые клетки находятся в эпителии перепончатого лабиринта внутреннего уха, где они образуют отдельные группы, или «пятна». Волосковые клетки каждой группы закреплены на месте с помощью промежуточных поддерживающих клеток, а сверху над ними нависает слой студенистого внеклеточного матрикса, соединенный с кончиками стереоцилий (рис. 19-46). При смещении слоя матрикса стереоцилий наклоняются, и в результате механической деформации волосковых клеток в них создается рецепторный потенциал (рис. 1947), Специфические функции различных групп волосковых клеток определяются главным образом природой структур, которые окружают эти клетки и через которые передается силовое воздействие. В случае волосковых клеток, реагирующих на линейное ускорение и силу тяжести, нависающий матрикс оттягивается вниз благодаря плотным кристаллам карбоната кальция: при ускоренном движении головы или ее наклоне матрикс смещается относительно волосковых клеток и стереоцилий сгибаются. В отличие от этого волосковые клетки, чувствительные к вращательному ускорению, расположены так, что при повороте головы на нависающий матрикс воздействует боковая сила, вызванная током жидкости в полукружных каналах внутренного уха.
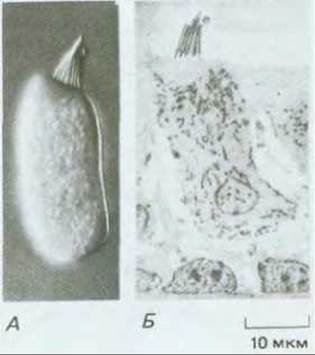
Рис. 19-45. А. Микрофотография сенсорной волосковой клетки, выделенной из внутреннего уха лягушки-быка. Можно видеть пучок стереоцилий, расположенный на верхушке клетки. Б. Микрофотография (полученная при малом увеличении с помощью трансмиссионного электронного микроскопа) волосковой клетки в естественном окружении опорных клеток. (A.J. Hudspeth, Science, 230, 745-752, 1985. Copyright 1985 by the AAAS.)
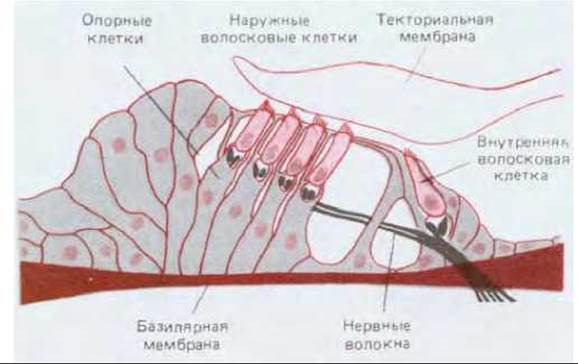
Рис. 19-46. Схематический поперечный разрез органа слуха (кортиева органа) во внутреннем ухе млекопитающих. Видно, что слуховые волосковые клетки находятся внутри сложных структур, образуемых опорными клетками; над слуховыми клетками нависает текториальная мембрана (слой внеклеточного матрикса). Полагают, что внутренние волосковые клетки ответственны в основном за слух - благодаря особому механизму преобразования, описанному в тексте. Эти клетки образуют синапсы с нейронами, передающими слуховые сигналы от уха к мозгу. В отличие от этого наружные волосковые клетки богато иннервированы дополнительной группой аксонов, передающих сигналы от мозга; функция этих клеток все еще загадочна. Имеются данные, позволяющие предполагать, что наружные волосковые клетки способны каким-то образом действовать как преобразователи в обратном направлении и составляют часть системы обратной связи, регулирующей передачу механических стимулов внутренним волосковым клеткам.
Волосковые клетки, улавливающие звук в ухе млекопитающего, образуют наиболее сложно организованную систему (рис. 19-46). Слуховые волосковые клетки располагаются рядами на базилярной мембране - узкой и длинной упругой перегородке между двумя заполненными жидкостью спиральными каналами, идущими параллельно в особом отделе внутреннего уха, называемом улиткой. Звуковые волны вызывают вибрацию барабанной перепонки, и через крошечные косточки среднего уха эта вибрация передается жидкости в каналах улитки и далее базилярной мембране, колебания которой заставляют стереоцилии слуховых волосковых клеток наклоняться. Благодаря особому устройству улитки, разные участки которой резонируют в разной степени в зависимости от частоты звуковых волн, распределение активируемых волосковых клеток доставляет информацию о высоте звука.
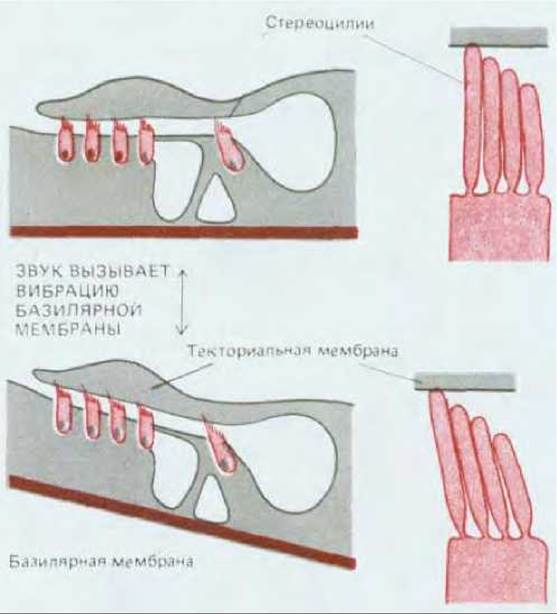
Рис. 19-47. Здесь показано, каким образом движение нависающего слоя внеклеточного матрикса (текториальной мембраны) отклоняет стереоцилии слуховых волосковых клеток во внутреннем ухе млекопитающих. Стереоцилии ведут себя как жесткие палочки с «шарнирным» прикреплением у основания. Верхушки пучков стереоцилии способны механически взаимодействовать с нависающим матриксом, либо непосредственно соприкасаясь с ним, либо через вязкую жидкость, находящуюся в промежутке между ними.
19.6.3. При наклоне пучков стереоцилий открываются механически регулируемые катионные каналы в их кончиках [40, 41]
При резком сдвиге слоя матрикса, нависающего над группой волосковых клеток, стереоцилий отклоняются в сторону на несколько градусов: при этом проницаемость клеточной мембраны изменяется и возникает направленный внутрь клетки ток, называемый рецепторным током (рис. 19-48). Величина ответа выходит на плато через 100-500 мкс, что соответствует времени, необходимому для открытия ацетилхолин-активируемого катионного канала в нервно-мышечном соединении, но гораздо меньше, чем нужно для возникновения электрических изменений при активации любого из известных рецепторов, не связанных с каналами. Поэтому кажется весьма вероятным, что механический стимул непосредственно открывает ионный канал. Как показали эксперименты с изменением внеклеточных концентраций ионов, механически регулируемый ионный канал, подобно рецептору ацетилхолина, практически одинаково проницаем для всех небольших катионов, и проходящий через него ток образуют главным образом ионы калия. (Ионная среда внутри уха несколько необычна, и на мембране волосковой клетки создается большой электрохимический градиент К+.) Но в какой части волосковой клетки находятся такие каналы и как преобразование сигнала связано со сложным строением клетки?
На каждой клетке стереоцилий располагаются плотными рядами разной высоты (подобно трубам в органе) (см. рис. 19-45 и 19-49). Позади середины самого высокого ряда часто имеется одна настоящая ресничка, или киноцилия, которая всегда присутствует в период развития, хотя не принимает участия в преобразовании сигнала и иногда потом исчезает (как у слуховых волосковых клеток млекопитающих). Если с помощью микрозонда отклонять стереоцилий, то они будут вести себя как упругие палочки, связанные в пучок; при этом каждая палочка поворачивается вокруг точки прикрепления к поверхности волосковой клетки и скользит относительно соседних палочек, так что и кончики стереоцилий смещаются относительно друг друга. Как показывает электронная микроскопия, помимо бокового сцепления, связывающего стереоцилий в пучок, имеются еще тонкие нити, направленные более или менее вертикально от верхушки каждой более короткой стереоцилий к расположенной выше точке на ближайшей более длинной стереоцилий (рис. 19-49). Как показали опыты с микроэлектродом, максимальная деполяризация мембраны волосковой клетки наблюдается при таком наклоне стереоцилий, когда тонкие вертикальные нити сильнее всего натянуты. По-видимому, трансмембранный ток, возникающий в результате наклона стереоцилий и приводящий к появлению рецепторного потенциала, поступает в волосковую клетку вблизи верхушки стереоцилий. Таким образом, вся структура в целом предназначена для того, чтобы ионный канал на кончике стереоцилий открывался в результате механического натяжения, вызываемого наклоном пучка стереоцилий (рис. 19-50).
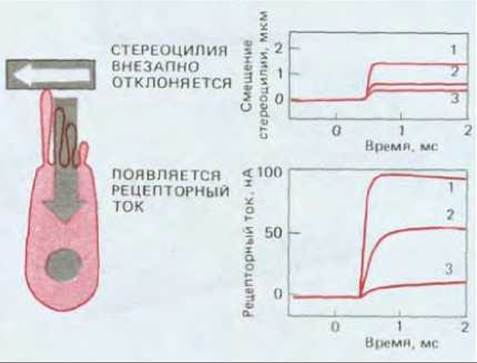
Рис. 19-48. Записи рецепторных токов (слева), входящих в волосковые клетки внутреннего уха лягушки-быка при внезапном отклонении пучков стереоцилий. Чем больше отклоняются стереоцилий, тем сильнее ток. (По данным D.P. Corey, A.J. Hudspeth, J. Neurosci., 3, 962-976, 1983.)
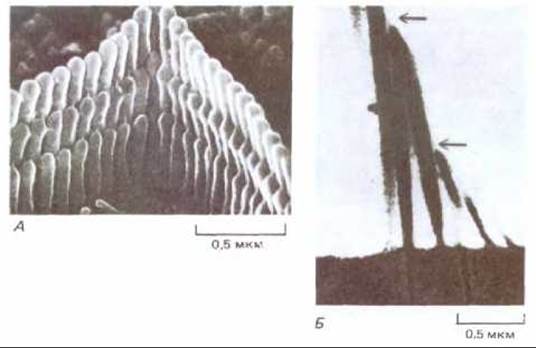
Рис. 19-49. А. Микрофотография слуховой волосковой клетки млекопитающего, полученная с помощью сканирующего электронного микроскопа. Видны тонкие филаменты, идущие от верхушек коротких стереоцилий к более длинным стереоцилиям следующего ряда. Б. Те же структуры, как они видны в трансмиссионном электронном микроскопе. Стрелками показаны филаменты. (Б- фотография М. P. Osborne; А и Б воспроизведены с разрешения J. D. Pickles, Prog. Neurobiol., 24, 1-42, 1985. Copyright 1985, Pergamon Press plc.)
Механизм волосковых клеток удивительно чувствителен: самые слабые звуки из тех, какие мы только можем слышать, растягивают вертикальные нити, прикрепленные к верхушкам стереоцилий, в среднем на 0,04 нм, что вдвое меньше диаметра атома водорода. Анализ рецепторного тока показывает, что в стереоцилий, вероятно, находится от одного до пяти механически регулируемых каналов. Каждая слуховая волосковая клетка человека несет около сотни стереоцилий, а в одном ухе имеется около 3500 таких клеток, так что способность слышать нам обеспечивают менее 4 млн. молекул-преобразователей.
19.6.4. Фоторецепторы обладают высокой чувствительностью и способны к адаптации, но реагируют относительно медленно [42]
Чувствительность фоторецепторов в глазах позвоночных приближается к крайнему пределу, допускаемому квантовой природой света. Кроме того, диапазон восприятия чрезвычайно широк - от света максимальной переносимой яркости до едва заметного освещения. Но по сравнению со слуховыми преобразователями скорость реагирования фоторецепторов очень мала. При оптимальных условиях самому «быстрому» фоторецептору человеческого глаза требуется примерно 25 мс для достижения пика электрической реакции на вспышку света, что более чем в 100 раз больше времени реагирования типичной волосковой клетки. Вероятно, относительная медленность ответа фоторецептора связана с важными ограничениями, заложенными в природе механизма зрительного преобразования.
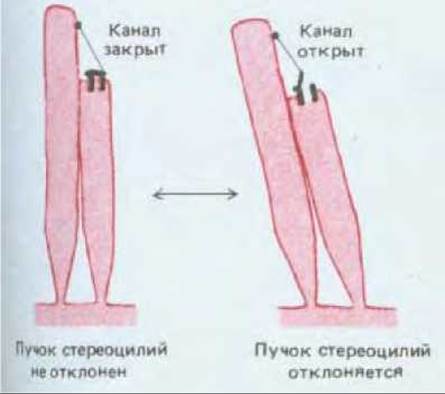
Рис. 19-50. Схема предполагаемого механизма, благодаря которому в результате наклона стереоцилий волосковой клетки открывается ионный канал. С помощью тонких механических измерений, подтверждаемых результатами регистрации электрического состояния одиночной волосковой клетки при отклонении пучка стереоцилий с помощью гибкого стеклянного зонда, можно действительно обнаружить ответ механически зависимых каналов на приложенную силу. С помощью этого метода можно показать, что сила, необходимая для открытия одного гипотетического канала, равна примерно 2∙10-13 ньютонов и что его «ворота» при этом перемещаются на расстояние около 4 нм.
19.6.5. Рецепторный потенциал, возникающий в палочке, - результат закрытия натриевых каналов [43]
Как уже говорилось в гл. 17 (разд. 17.2.2), в глазу позвоночных имеются два типа фоторецепторных клеток. Колбочки служат для цветового зрения и восприятия мелких деталей и требуют сравнительно сильной освещенности. Палочки обеспечивают монохроматическое зрение при слабом свете и могут дать измеримый электрический ответ на один-единственный фотон (рис. 19-51). Механизм действия палочек и колбочек, по-видимому, сходен, но палочки лучше изучены.
Палочка (рис. 19-52) состоит из наружного сегмента, содержащего световоспринимающий аппарат, внутреннего сегмента, где находится множество митохондрий, ядерной области и (в основании клетки) синаптического тельца, образующего контакт с нервными клетками сетчатки (см. рис. 17-6). Как это ни удивительно, но в темноте клетка очень сильно деполяризована; эта деполяризация удерживает потенциал-зависимые кальциевые каналы синаптического тельца в открытом состоянии, и переход ионов Са2+ внутрь клетки приводит к непрерывному высвобождению медиатора. Деполяризация обусловлена тем, что в плазматической мембране наружного сегмента открыты натриевые каналы. При воздействии света эти каналы закрываются, так что рецепторный потенциал проявляется в форме гиперполяризации, приводящей к снижению притока Са2+ и уменьшению скорости выделения медиатора (рис. 19-52). Так как медиатор оказывает тормозящее действие на многие из постсинаптических нейронов, эти нейроны при освещении растормаживаются и в результате возбуждаются. Скорость высвобождения медиатора фоторецепторами изменяется в соответствии с интенсивностью света: чем ярче свет, тем сильнее гиперполяризация и тем больше замедляется выделение медиатора. При очень слабом фоновом освещении, когда клетка находится в наиболее чувствительном, «темноадаптированном» состоянии, поглощение одного фотона снижает приток натрия на миллион или более ионов Na+, и гиперполяризация достигает ~1 мВ.
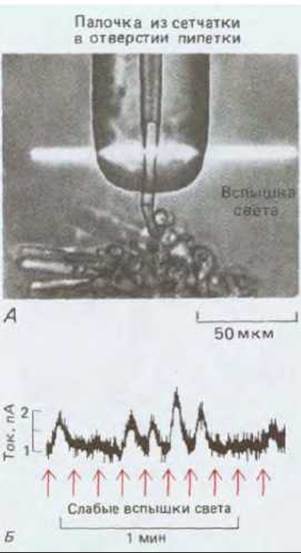
Рис. 19-51. Электрическая реакция палочки на одиночные фотоны. А. На микрофотографии показана методика проведения эксперимента. Отпрепаровывают кусочек сетчатки жабы, и наружный сегмент одной палочки всасывают в отверстие стеклянной микро-пипетки, которая затем служит электродом для регистрации тока, проходящего через мембрану палочки. Б. Запись изменений силы тока во время серии слабых вспышек света; число фотонов, поглощаемых клеткой при каждой вспышке, варьирует случайным образом, но оно всегда целое. Высокие пики на графике чаще всего соответствуют поглощению одного или двух фотонов, но многие вспышки не вызывают никакой реакции, так как поглощения фотонов не происходит. (D. A. Baylor et al, J. Physiol., 288, 589-611, 1979.)
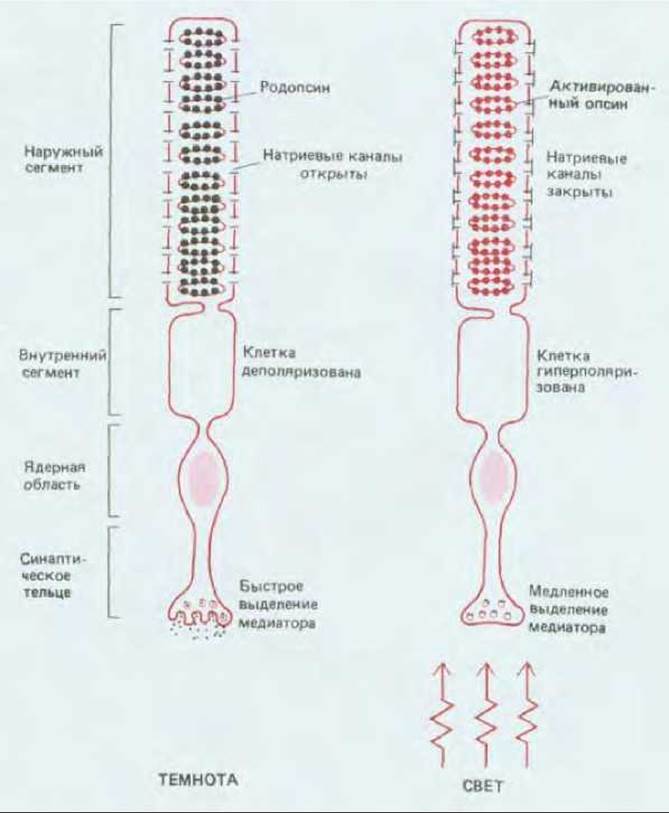
Рис. 19-52. Реакция палочки на освещение. Фотоны поглощаются случайным образом молекулами родопсина, находящимися в наружном сегменте. Это приводит к закрытию натриевых каналов в плазматической мембране и снижению скорости выделения медиатора из синаптического тельца.
19.6.6. Фотоны изменяют конформацию молекул родопсина [43, 44]
Каким образом свет первоначально воспринимается клеткой и какова цепь событий, приводящих к закрытию натриевых каналов? Наружный сегмент, где происходят ключевые этапы преобразования светового сигнала, представляет собой цилиндр, содержащий около тысячи дисков, плотно упакованных в виде стопки (см. рис. 17-7). Каждый диск образован замктутой в пузырек мембраной, в которой находятся светочувствительные молекулы родопсина; плотность упаковки этих молекул составляет примерно 105 на 1 мкм2. Молекула родопсина состоит из трансмембранного гликопротеина опсина (длиной 348 аминокислотных остатков) и ковалентно связанной с ним простетической группы 11 -цис-ретиналя, который и поглощает свет. При поглощении фотона 11-цис-ретиналь почти мгновенно изомеризуется в полностью-транс-ретиналь, изменяя при этом свою форму, что приводит к более медленному изменению конформации опсина. Все эти события занимают около 1 мс. Затем примерно через 1 мин полностью-транс-ретиналь отделяется от опсина в результате гидролиза связи между ними и выходит в цитозоль, где в конце концов снова переходит в 1 1-цис-форму; последняя соединяется с опсином, и таким образом происходит регенерация светочувствительной молекулы родопсина. Именно раннее конформационное изменение родопсина, вызванное светом, приводит к закрытию натриевых каналов. Но поскольку родопсин находится в дисках, т. е. на некотором расстоянии от каналов, для сопряжения этих событий необходим «посредник».
19.6.7. К закрытию натриевых каналов плазматической мембраны приводит снижение уровня циклического GMP в цитоплазме фоторецепторных клеток, вызванное светом [43, 44, 45]
При улавливании света палочкой происходит изменение внутриклеточной концентрации как Са2+, так и cGMP, поэтому любая из этих молекул могла бы в принципе служить внутриклеточным посредником. С помощью метода исследования небольших участков мембраны (разд. 4.2.3) установлено, что ключевым сигналом служит падение концентрации циклического GMP в цитозоле. В решающем эксперименте небольшой участок мембраны наружного сегмента отсасывали с помощью микроэлектрода, и тогда внутренняя сторона этого участка становилась доступной для воздействий (рис. 19-53). Когда в среду, омывающую кусочек мембраны, вводили циклический GMP, а к мембране прикладывали разность электрохимических потенциалов, появлялся ток ионов Na+ после удаления циклического GMP ток прекращался, несмотря на высокую концентрацию Са2+. Таким образом, сGМР открывает натриевые каналы, а свет, вызывающий снижение концентрации cGMP, закрывает эти каналы. Обычно эффекты циклических нуклеотидов реализуются через фосфорилирование определенных белков активированной протеинкиназой (разд. 12.3.14), однако в палочках cGMP воздействует прямо на натриевые каналы, не позволяя им закрываться. Но каким образом вызванное светом изменение конформации родопсина снижает концентрацию cGMP в цитозоле палочки?
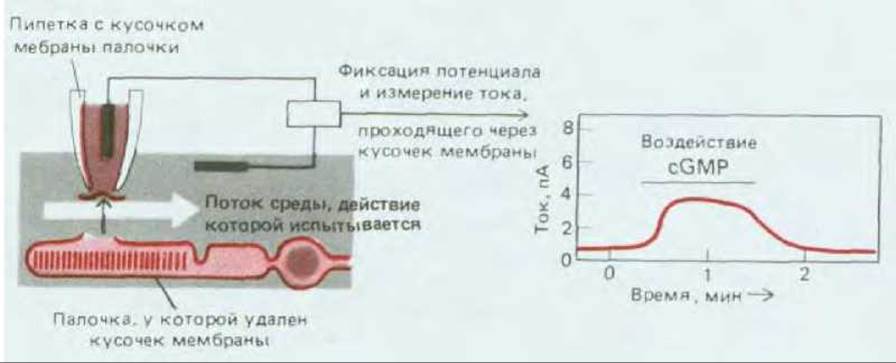
Рис. 19-53. Схема эксперимента, который показывает, что cGMP при его воздействии на плазматическую мембрану палочки со стороны цитоплазмы непосредственно контролирует открытие и закрытие ионных каналов. Относительно медленное увеличение и уменьшение тока при добавлении и удалении cGMP связано с тем, что для изменения состава омывающей среды требуется некоторое время. Скорость реакции каналов на cGMP слишком высока, чтобы ее можно было определить с помощью этой методики. (По Е. Е. Fesenko, S. S. Kolesnikov, A. L. Lyubarsky, Nature, 313, 310-313, 1985.)
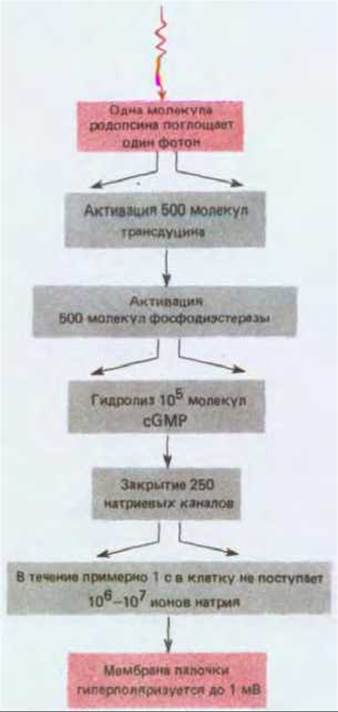
Рис. 19-54. Каскад ферментативных реакций, приводящий к возникновению рецепторного потенциала после поглощения одного фотона палочкой, адаптированной к темноте. Расходящимися стрелками указаны этапы, на которых происходит усиление.
Поглощение одного фотона одной молекулой родопсина приводит к гидролизу многих молекул cGMP. Такое усиление эффекта достигается благодаря каскаду ферментативных реакций. Одна молекула активированного родопсина катализирует активацию G-белка, называемого трансдуцином, с очень высокой скоростью - примерно 1000 молекул в секунду. Трансдуцин гомологичен Gs-белку (разд. 12.3.4), который функционально сопрягает рецепторы с аденилатциклазой (а сам родопсин гомологичен таким рецепторам - см. разд. 12.3.12). Однако активированный трансдуцин не взаимодействует с аденилатциклазой, а активирует cGMP-фосфодиэстеразу, которая специфически гидролизует cGMP со скоростью около 4000 молекул в секунду, что приводит к быстрому снижению уровня cGMP. В результате всего этого каскада, занимающего примерно секунду, гидролизуется более 105 молекул cGMP на один поглощенный квант света, что приводит к кратковременному закрытию 250 натриевых каналов (рис. 19-54).
19.6.8. Фоторецептор адаптируется к яркости света [46]
Для возвращения фоторецептора в состояние покоя после возбуждения его светом каждая реакция ферментативного каскада, инициированного светом, должна быть «уравновешена» соответствующей реакцией инактивации. По-видимому, свет ускоряет как активирующие, так и инактивирующие реакции, но второй эффект проявляется чуть позднее, поэтому свет вызывает мгновенный положительный ответ, который затем очень быстро затухает. Такая запаздывающая инактивация не только помогает обеспечить короткий ответ на короткую вспышку света, но и дает возможность фоторецептору адаптироваться: свет постоянной яркости, вместо того чтобы просто приводить клетку в состояние насыщения с близкой к нулю концентрацией cGMP, вызывает два противоположных эффекта, которые почти гасят друг друга, что позволяет клетке реагировать на последующие изменения освещенности.
По-видимому, вызванное светом уменьшение концентрации Са2+ играет решающую роль как в прекращении реакции на вспышку света, так и в адаптации. Если изменение концентрации Са2+ искусственно задержать путем введения в фоторецептор Са2+-буфера, то электрический ответ на световую вспышку будет очень продолжительным и клетка будет слишком медленно адаптироваться к постоянному освещению. Если фоторецептор поместить в раствор, полностью блокирующий передвижение Са2+ через плазматическую мембрану, то те же самые эффекты проявятся в еще большей степени, а адаптации не произойдет совсем. В норме каналы, через которые ионы натрия поступают в наружный сегмент фоторецептора, до некоторой степени проницаемы и для других катионов, включая Са2+. Свет, закрывая каналы, блокирует приток кальция, в то время как отток Са2+ (опосредуемый в плазматической мембране палочки системой Са2+/Nа+-антипорта) продолжается, в результате чего внутриклеточная концентрация ионов кальция падает.
Полагают, что это ускоряет те ферментативные реакции (в особенности синтез cGMP гуанилатциклазой), которые противодействуют вызываемому светом падению концентрации cGMP, помогая клетке адаптироваться.
19.6.9. Нейроны обрабатывают исходную информацию, доставляемую сенсорными рецепторными клетками [47]
Через сенсорные рецепторные клетки в нервную систему поступает огромный поток информации. Мозг должен переработать эту информацию и выделить значимые элементы: выхватить слова из хаоса звуков, различить лицо среди светлых и темных пятен и так далее. В этом заключается вторая стадия переработки сенсорной информации - переработка на уровне нейронов, гораздо более тонкая и сложная, чем та, что происходит в рецепторных клетках. Эта вторая стадия включает «вычисления», выполняемые сложно переплетенной сетью нейронов, где каждый нейрон обычно получает множество сигналов, среди которых есть и возбуждающие, и тормозные. Каждый нейрон генерирует выходной сигнал, несущий информацию о наличии или отсутствии каких-то специфических элементов в исходных данных, доставляемых рецепторными клетками. Например, определенные группы клеток в зрительных центрах мозга генерируют потенциалы действия, когда глаз видит прямую линию, определенным образом ориентированную в пространстве. Выходные сигналы от одной группы будут приняты другими нейронами, выполняющими следующий этап процесса, и так далее ко все более высоким уровням восприятия - вплоть до распознавания таких тонких и сложных вещей, как осмысленные слова и выражения лиц.
Для такой переработки информации необходима поразительно сложная организация анатомических связей между нервными клетками. Детали взаимоотношений между анатомией нервной системы и высшей нервной деятельностью выходят за рамки книги о биологии клетки. Но в чем все же заключаются основные механизмы, благодаря которым создаются сложные, но упорядоченные анатомические структуры? Рассмотрению этого вопроса будет посвящен следующий раздел главы.
Заключение
Специальные преобразователи превращают сенсорные стимулы в электрические сигналы. Например, у позвоночных волосковые клетки внутреннего уха представляют собой механорецепторы: на свободной поверхности каждой волосковой клетки имеется пучок стереоцилий (гигантских микроворсинок), и при наклоне таких пучков открываются ионные каналы, что ведет к изменению мембранного потенциала. Мембранный потенциал фоторецепторных клеток в глазу позвоночного изменяется при поглощении света молекулами родопсина, содержащимися в этих клетках. И в том, и в другом случае электрический сигнал, возникающий в сенсорной клетке вначале в форме рецепторного потенциала, передается соседним нейронам через химические синапсы. Однако два упомянутых класса клеток-рецепторов используют для выработки рецепторных потенциалов различные «стратегии»: в основе одной лежат рецепторные молекулы, связанные с каналами, а другая зависит от молекул-рецепторов, не связанных с каналами. В волосковых клетках физическая связь между стереоцилиями порождает механические силы, которые прямо воздействуют на ионные каналы в плазматической мембране, заставляя их быстро открываться или закрываться. В палочках сетчатки активированные светом молекулы родопсина инициируют каскад ферментативных реакций, в результате которых в цитозоле гидролизуется циклический GMP, что в свою очередь ведет к закрытию натриевых каналов плазматической мембраны. Хотя механизм, основанный на каталитических реакциях, не может быть очень быстрым, он позволяет реагировать на один-единственный фотон.