Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Особенности растительных клеток
Взаимодействия между растениями и другими организмами
И корни, и побеги растений взаимодействуют с бесчисленным множеством организмов, среди которых бактерии, грибы, черви и насекомые. Для этого взаимодействия в ходе эволюции выработались специальные механизмы (генетические, химические и анатомические). К ним относятся защитные реакции против патогенов и симбиотические связи с организмами, необходимые для жизни растения. В данном разделе мы рассмотрим четыре примера подобных взаимодействий. Они иллюстрируют всю сложность возникающих при этом сигналов и ответов на них.
20.3.1. Большинство сосудистых растений живет в симбиозе с почвенными грибами [14]
Если проростки лесного дерева, выращенные в стерильных условиях, высадить в луговую почву, то скорее всего они погибнут. Однако если проростки высадить в землю, куда добавлено небольшое количество лесной почвы, то нормальный рост будет обеспечен (рис. 2028). Необходимый фактор, содержащийся в лесной почве, - это грибы, которые вступают в тесную симбиотическую связь с корнями растения и образуют микоризу или «грибной корень», обнаруживаемый более чем у 90% сосудистых растений. Нити грибницы обладают очень большой поверхностью и образуют покрытие корня растения, напоминающее войлок; при этом они проникают между, а в некоторых случаях и внутрь клеток коры корня. Грибы секретируют факторы роста, заставляющие клетки корневой коры увеличиваться в размерах и делиться, что обусловливает ветвление корней. Изменяя рН почвы, грибы также повышают содержание в ней неорганических питательных веществ, в особенности фосфатов, переводя их в форму, которая может усваиваться растением. В свою очередь растение снабжает грибы сахарами и аминокислотами. Каждое растение образует микоризу лишь с ограниченным числом видов грибов, и во многих случаях грибы необходимы для выживания растений. Например, хороший рост орхидей после прорастания семян полностью зависит от присутствия соответствующих грибов. Не исключено, что географическое распространение многих растений определяется присутствием грибов, связанных с ними. Молекулярные механизмы взаимодействия гриба и растения в микоризе еще неясны. Значительно больше известно о симбиозе корней растений с бактериями.
20.3.2. Бактерии-симбионты помогают некоторым растениям усваивать атмосферный азот [15]
Исходные материалы, необходимые для роста растений, представлены главным образом неорганическими веществами - О2, СО2, Н2О и солями. Большинство солей, потребляемых корнями из почвы, образуются в результате разрушения горных пород. Исключение составляет азот: весь, содержащийся в живых организмах азот происходит в конечном счете из атмосферного азота, который включается в органические вещества в ходе весьма энергоемкого процесса (с этим связана и высокая цена искусственно производимых азотосодержащих удобрений). Единственными организмами, способными фиксировать азот из атмосферы, являются прокариоты (цианобактерии и отдельные виды группы эубактерий). Некоторые из них - свободноживущие почвенные организмы, другие же (например, бактерия Rhizobium) вступают в симбиотическую ассоциацию с корнями определенных растений, например бобовых - гороха, бобов и клевера.

Рис. 20-28. Воздействие микоризы на рост растений. Все проростки сосны первые два месяца выращивали в стерильном растворе питательных веществ (здесь они изображены в возрасте 9 месяцев). Одни проростки (А) высаживали сразу в почву, взятую из открытой степи, а другие (Б) перед перенесением в степную почву две недели выращивали в почве, взятой из леса. Эти последние (Б) растения имели преимущество в росте и развитии благодаря наличию в лесной почве определенныхгрибов, образующих микоризу. (С любезного разрешения J. lyer, S. Wilde.)
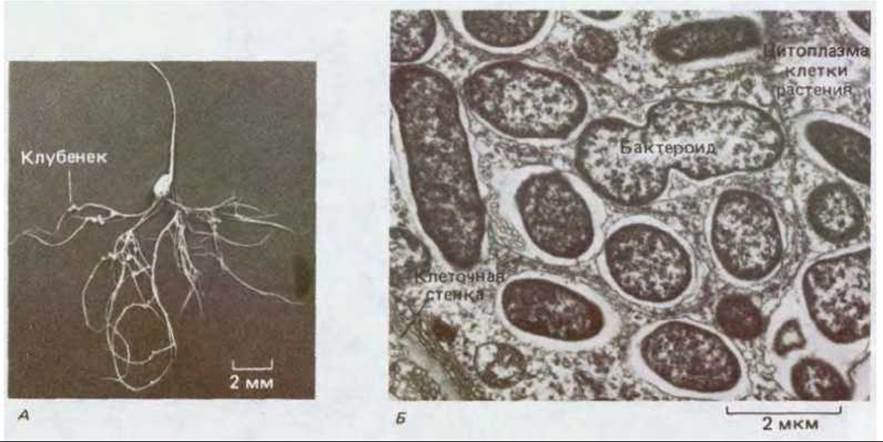
Рис. 20-29. А. Молодой проросток гороха в симбиотической ассоциации с азотфиксирующими бактериями Rhizobium. Хорошо видны корневые клубеньки, в которых находятся бактерии. Б. Электронная микрофотография тонкого среза корневого клубенька гороха, показанного на рис. А. Азотфиксирующие бактероиды Rhizobium, окруженные мембраной, принадлежащей клетке-хозяину, заполняют цитоплазму этой клетки. (А - с любезного разрешения A. Johnston; Б - с любезного разрешения В. Huang, Q. S. Ma.)
Первая стадия в возникновении симбиоза - специфическое узнавание бактерией тонких корневых волосков, отходящих от специализированных эпидермальных клеток растения-хозяина. После связывания с клетками эпидермиса корня растущая бактерия проникает в растение с помощью инфекционных филаментов и вызывает деление кортикальных клеток, лежащих под эпидермальными; в результате образуется большой корневой клубенек (рис. 20-29, А). Бактерии внедряются во все новые кортикальные клетки, заселяя их цитоплазму. Примерно половина массы каждого зрелого клубенька приходится на внутриклеточные бактерии, которые утратили большую часть своей клеточной стенки. Плазматическая мембрана каждой такой бактерии окружена еще одной мембраной, которую продуцирует клетка-хозяин. Именно эти видоизмененные бактерии, именуемые бактероидами, и фиксируют азот, который в конечном итоге используется растением (рис. 20-29, Б).
Бактериальный фермент, катализирующий связывание азота, носит название нитрогеназа. Он представляет собой сложную белковую молекулу, состоящую из трех полипептидных цепей. У симбиотических видов Rhizobium этот белковый комплекс катализирует образование аммиака из атмосферного азота. Затем аммиак быстро перемещается в цитоплазму клеток хозяина, где он превращается в глутамин. В конечном счете фиксированный азот включается во все остальные аминокислоты.
Генетический анализ показал, что для возникновения и поддержания подобного симбиоза необходима координированная экспрессия многих генов, принадлежащих и бактерии, и растению. «Диалог» между бактерией и клеткой-хозяином начинается со связывания бактерии с корневыми волосками. В результате этого связывания активируется ряд генов хозяина, кодирующих в растительной клетке белки (нодулины), которые необходимы для роста и функционирования клубенька. Флавоноид, вырабатываемый клетками растения, связывается с белком, кодируемым бактериальным геном nod, и активирует его (рис. 20-30). Активированный белок nodD включает синтез продукта другого бактериального гена, который индуцирует образование нодулинов растения-хозяина. Большая часть бактериальных генов nod, а также гены nif (они кодируют белки, участвующие в фиксации азота, например нитрогеназу) содержатся в большой плазмиде. Если плазмидные гены азотфиксации, содержащиеся у Rhizobium, образующей клубеньки на фасоли, заменить на соответствующие гены, выделенные из клеток штамма, специфичного для гороха, бактерии начинают формировать клубеньки только на горохе.
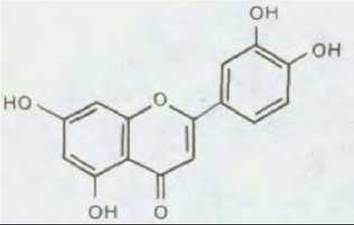
Рис. 20-30. Различные флавоны и родственные им соединения, выделяемые корнем растения-хозяина, связываются со специфическими бактериями, образующими клубеньки, и активируют их. На рисунке представлена структура лютеолина флавона люцерны, который у Rhizobium meliloti индуцирует гены nod. Эта сигнальная молекула, очевидно, связывается с белком nodD, который в активированной форме включает бактериальные гены азотфиксации. Флавоны тесно связаны с антоциановыми пигментами цветков и плодов. Различные растения продуцируют разные комбинации флавонов, которые избирательно активируют специфичные для этих растений виды Rhizobium.
К нодулинам клетки-хозяина относятся белки, участвующие в делении кортикальных клеток корня, структурные компоненты клубенька, ферменты дающие возможность растению ассимилировать вещества, содержащие связанный азот, и особые белки, необходимые для функционирования бактероида. Наиболее важный из этих белков - леггемоглобин - локализован в цитоплазме, связывающей кислород, и аналогичен миоглобину млекопитающих. Нитрогеназный комплекс бактерий необратимо инактивируется свободным кислородом. В связи с этим необходимы специальные механизмы, которые могли бы обеспечивать существование бактероида в корнях. Благодаря им давление кислорода поддерживается на таком уровне, чтобы обеспечить дыхание и вместе с тем не повредить нитрогеназу. Индуцируемый Rhizobium синтез больших количеств леггемоглобина, действующего как кислородный буфер, является частью этого механизма. Глобиновая часть молекулы кодируется геном хозяина, а гем (простетическая группа) детерминируется бактериальным партнером - замечательный пример эволюционной адаптации организмов друг к другу.
Процесс азотфиксации обеспечивается солнечной энергией, преобразованной посредством фотосинтеза. Установлено, что для фиксации одной молекулы азота Rhizobium необходимо около 25-35 молекул АТР. Эти прокариоты поставляют для растений в естественных экосистемах гораздо больше фиксированного азота (~2 х 108 тонн в год), чем можно получить при использовании азотных удобрений.
20.3.3. Agrobacterium представляет собой фитопатоген, переносящий гены в геном своего хозяина [16]
Другая почвенная бактерия, близкородственная Rhizobium, называется Agrobacterium tumefaciens. Эта бактерия у растений вызывает образование корончатых галлов. При контакте с Agrobacterium нормальные растительные клетки трансформируются в опухолевые, вследствие переноса генов от бактерии к хозяину (рис. 20-31).
Опухолевые клетки, индуцируемые Agrobacterium, обладают рядом замечательных свойств. Во-первых, такие клетки можно выделить и неограниченно долго выращивать в культуре без факторов роста и вне контакта с клетками Agrobacterium. Во-вторых, они синтезируют ряд необычных веществ, называемых опинами. Эти производные аминокислот могут перерабатываться и усваиваться преимущественно тем штаммом бактерий, которые индуцировали синтез опинов в растительных клетках.
Многие свойства опухолевых клеток стали понятны при анализе молекулярно-генетических данных. Способность Agrobacterium индуцировать опухоли связана с большой ДНК-содержащей плазмидой, названной Ti (от англ. tumor inducing - вызывающая опухоли). Часть этой плазмиды-Т-ДНК (от англ. transferred ЭКА-передаваемая ДНК) включается в ядерный геном растительной клетки. Бактерия может проникнуть в чувствительное растение только в том месте, где оно повреждено, т. е. там, где растительные клетки секретируют необычные фенольные соединения, в частности ацетосирингон (рис. 20-32). При этом у бактерии запускается каскад реакций, в результате которых Т-ДНК вырезается из Ti-плазмиды и переносится в геном клетки-хозяина (рис. 20-33). Включившись в хромосому клетки-хозяина, Т-ДНК транскрибируется и транслируется в ней, образуя три класса белков.

Рис. 20-31. Опухоли, индуцированные Agrobacterium tumefaciens у комнатного растения-суккулента. (С любезного разрешения P. Hooykaas, из Genetic Eng., I, 155, 1979.)
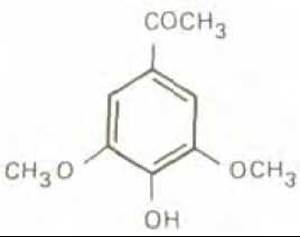
Рис. 20-32. Ацетосирингон сигнальная молекула, обнаруженная в поврежденных, но метаболически активных клетках растений. Ацетосирингон специфически активирует вирулентные гены Ti-плазмиды Agrobacterium, которые принимают участие в образовании Т-ДНК, интегрирующейся с геномом клетки-хозяина (см. рис. 20-33).
К первому относится фермент, вызывающий у растения синтез специфического опина, а два других - это ферменты, катализирующие синтез регуляторов роста растений - индолилуксусной кислоты и цитокинина (см. рис. 20-67). Повышение содержания этих двух регуляторов роста в результате активности включившихся в геном растения генов Т-ДНК вызывают неограниченный рост и деление трансформированных растительных клеток, чем и объясняется способность этих клеток продолжать рост в отсутствие обоих регуляторов роста и исходной бактерии.
Способность Т-ДНК Agrobacterium стабильно включаться в геном хозяина дает возможность широко использовать полученные на ее основе рекомбинантные молекулы ДНК в качестве вектора для генетической трансформации растительных клеток (см. разд. 20.5.10).
20.3.4. Продукты, возникающие при разрушении клеточной стенки, часто используются в качестве сигналов при взаимодействии растения и патогена [17]
В качестве защитных агентов растения используют большое число крайне специализированных метаболитов. Некоторые вещества, как, например, гликозиды в горчичном масле и самые разнообразные алкалоиды (кофеин, морфин, стрихнин и колхицин) действуют отпугивающе на травоядных животных. Кроме этих конститутивных средств защиты растения выработали более сложные адаптационные механизмы, которые приводятся в действие только при взаимодействии хозяина и патогена.
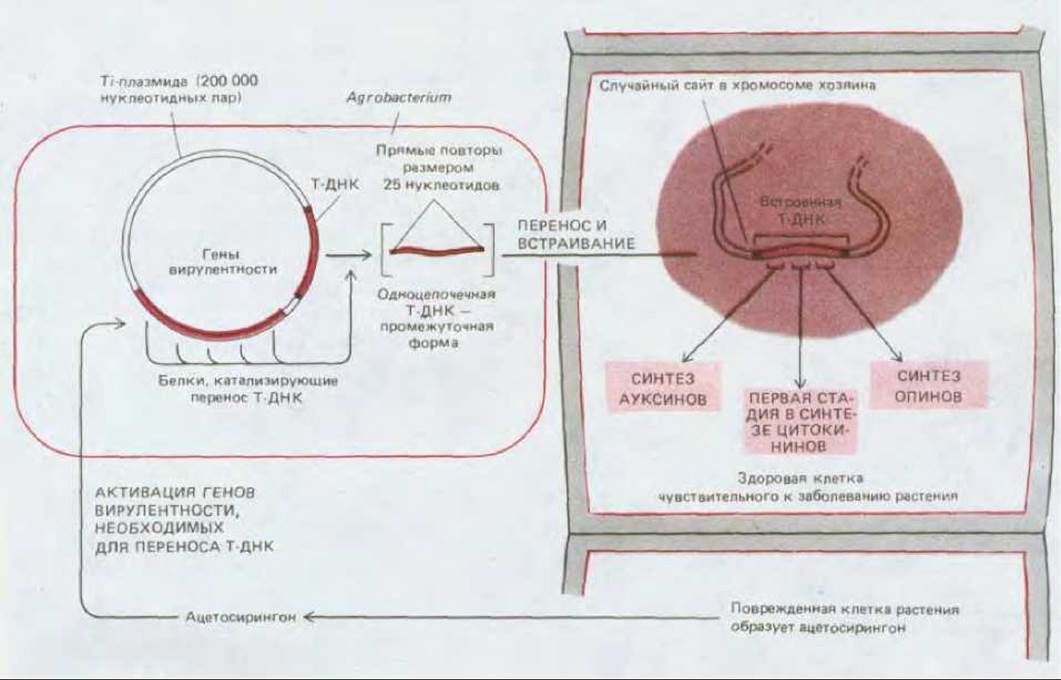
Рис. 20-33. Некоторые события, имеющие место при заражении растения бактериями Agrobacterium tumefaciens. Т-ДНК- участок Ti (от англ. tumor inducing - индуцирующей опухоли) - плазмиды бактерии встраивается в случайный сайт хромосомы растительной клетки-хозяина. Среди продуктов экспрессии ее генов присутствуют ферменты, участвующие в синтезе регуляторов роста растений, которые и приводят к возникновению опухоли.
Многие такие реакции протекают с участием компонентов клеточной стенки растения-хозяина или патогена. Например, в ответ на заражение бактерией или грибом растительные клетки укрепляют свои стенки за счет дополнительного отложения полимеров, входящих в их состав, включая лигнин и гликопротеины, обогащенные гидроксипролином. В свою очередь у патогенов в их войне с растением есть собственное оружие. Рассмотрим, например, что происходит, когда патогенный гриб (Fmarium solani) «атакует» растение гороха. Споры гриба секретируют очень небольшие количества фермента кутиназы, которая вызывает частичный гидролиз кутикулы, покрывающей лист. Высвободившиеся мономеры кутина стимулируют усиленное образование кутиназы спорой гриба. В результате кутикула растворяется, и прорастающие гифы гриба могут проникнуть в лист.
Однако продукты распада клеточной стенки, стимулируемые вторгающимся патогеном, могут оказаться полезными в качестве самых первых сигналов об опасности, грозящей клеткам растения-хозяина. Клетки, контактирующие с патогеном, обычно синтезируют низкомолекулярные продукты, которые называются фитоалексинами и представляют собой антибиотики, токсичные для определенных патогенных бактерий и грибов. В настоящее время идентифицированы некоторые соединения, ответственные за стимуляцию биосинтеза фитоалексинов растением. Эти вещества, называемые элиситорами, представляют собой короткоцепочечные олигосахариды, образующиеся из полисахаридов клеточной стенки и проявляющие активность при очень низких концентрациях (109- 10-10 М). Одним из первых хорошо охарактеризованных элиситоров является гепта-β-глюкозид, выделяющийся из клеточной стенки гриба, поражающего сою (рис. 20-34). Олигосахаридные элиситоры синтеза фитоалексинов могут также продуцироваться клеточной стенкой растений. В этом случае они представляют собой фрагменты пектинового скелета, построенного из остатков галактуроновой кислоты, которые высвобождаются из клеточной стенки растения при действии ферментов, секретируемых либо внедряющимся патогеном, либо в некоторых случаях ферментами растительной клетки, активированными при повреждении.
Простое механическое повреждение растений может вызвать синтез белков, ингибирующих протеазы как насекомых, так и микроорганизмов (ингибиторы протеаз). Полагают, что подобная нормальная защитная реакция может локально индуцироваться небольшими фрагментами полисахаридов, входящих в состав пектина, и высвобождающихся при повреждении. По-видимому, существуют и пока не идентифицированные сигнальные механизмы, действующие на расстоянии, поскольку листья, удаленные от поврежденного листа, также начинают продуцировать ингиторы протеаз.
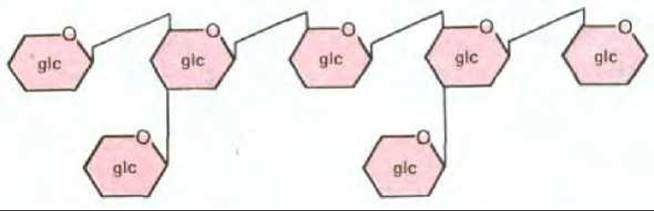
Рис. 20-34. Этот гептаглюкозид (олигосахарид, содержащий семь остатков глюкозы [glc]) выделяется клеточной стенкой Phytophthora megasperma - гриба, патогенного для сои. Данный олигосахарид имеет глюкозидный скелет (со связями ß1 → 6) и две боковые цепи (со связями ß1 → 3). При очень низких концентрациях гептаглюкозид специфически активирует ряд генов сои, включая те, которые принимают участие в биосинтезе фитоалексинов. Изменение связей или расположения любого из остатков глюкозы приводит к потере активности.
20.3.5. В нормальной растительной клетке клеточная стенка может быть важным источником сигналов [18]
Данные ряда экспериментов указывают на то, что сигналы, посылаемые клеткой, зависят от высвобождения олигосахаридов из клеточной стенки растений под действием ферментов. Такие события имеют место и при нормальном росте и развитии растения. Например, удлинение клетки ускоряется регулятором роста растений - ауксином (см. разд. 20.5.8), однако его воздействие может быть устранено даже очень низкими концентрациями сложного фрагмента гемицеллюлозы. Образование таких фрагментов из девяти остатков сахара стимулируется ауксином и, можно полагать, является составной частью регуляторной системы с обратной связью, которая в норме контролирует увеличение размеров клетки (см. разд. 20.4.7). Весьма вероятно, что клеточная стенка представляет собой место связывания и хранения многих сигнальных молекул, которые способны локально высвобождаться и действовать на развитие клетки. Эта гипотеза в настоящее время проверяется во многих лабораториях.
Заключение
В ходе развития растения неизбежно сталкиваются с самыми разнообразными бактериями и грибами. Многие из таких контактов полезны для растений. Так, например, у большинства из них корневые системы сосуществуют со сложными мицелиями грибов, благодаря которым питательные вещества становятся более доступными как для одного, так и для другого организма. Кроме того, почти весь азот, содержащийся в связанном виде в живых организмах, происходит в конечном итоге из азота атмосферы; азот воздуха фиксируется прокариотами, многие из которых (например, Rhizobium) образуют сложные симбиотические ассоциации с корнями растений. Но характер взаимодействия может быть и иным, полезным только для одного из организмов. Фитопатоген Agrobacterium вызывает образование опухолей на растении, вводя свою Т-ДНК в геном клетки-хозяина. Экспрессия вновь индуцированных генов приводит к возникновению дисбаланса молекул- регуляторов роста клеток растений. Специфические низкомолекулярные вещества, выделяемые растением, и у Rhizobium, и у Agrobacterium, активируют гены, необходимые для того, чтобы эти бактерии могли внедриться в растение. Другие небольшие молекулы действуют как своеобразные сигналы, которые запускают ответ хозяина на действие патогенов. В роли таких сигнальных молекул выступают специфические олигосахариды - продукты распада полисахаридов, входящих в состав клеточной стенки. Аналогичные соединения могут служить сигналами и при нормальном развитии растений.