Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Компоненты биомолекулярных комплексов
Липиды и биомембраны
Липидами называют природные соединения, которые получают из растительных или животных тканей экстракцией неполярными растворителями (например, эфиром, бензолом или хлороформом) и которые не растворимы в воде.
К ним относятся продукты взаимодействия жирных кислот со спиртами (простые липиды), аминоспиртами и другими соединениями (сложные липиды), простагландины и изопреноидные липиды (например, каротиноиды, хлорофилл, витамины Е и К).
Триацилглицеролы - простые липиды, представляющие собой эфиры трёхосновного спирта глицерола и трех жирных кислот (рисунок 55(a)). Они бывают различных типов в зависимости от природы трех остатков жирных кислот, присоединенных к гидроксильным группам глицерина.
Жирные кислоты также являются основой фосфолипидов, из которых образуются плазматические мембраны. Жирные кислоты состоят из углеводородной цепи и карбоксильной группы СООН. Они различаются по длине, хотя преимущественно в клетках жирные кислоты имеют четное число атомов углерода, обычно 14, 16, 18 и 20. Основные жирные кислоты фосфолипидов приведены в таблице 2.
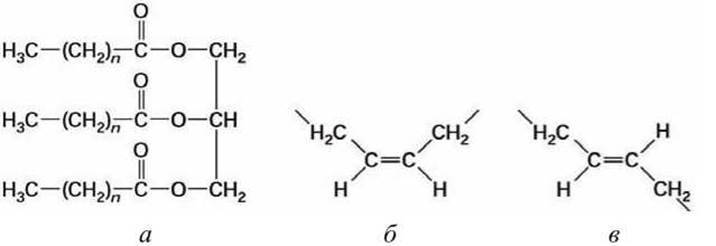
Рисунок 55 - Схема ориентации связей в липидах: а - триацилглицерол; б - цис-, в - транс-стереоизомерные конфигурации относительно двойной С=С связи
Жирные кислоты обычно обозначают Сх:у, где х - число атомов углерода в цепи, у - число двойных связей. Жирные кислоты, в которых нет двойных С=С связей, называют насыщенными, в случае, если есть хоть одна двойная связь, их называют ненасыщенными.
Таблица 2 - Основные жирные кислоты фосфолипидов
|
Название кислоты |
Обозначение |
Строение (химическая формула) |
|
Насыщенные жирные кислоты |
||
|
Миристиновая |
С14:0 |
СН3(СН2)12СООН |
|
Пальмитиновая |
С16:0 |
СН3(СН2)14СООН |
|
Стеариновая |
С18:0 |
СН3(СН2)16СООН |
|
Ненасыщенные жирные кислоты |
||
|
Олеиновая |
С18:1 |
СН3(СН2)7СН=СН(СН2)7СООН |
|
Линолевая |
С18:2 |
СН3(СН2)4СН=СНСН2СН=СН(СН2)7СООН |
|
Арахидоновая |
С20:4 |
СН3(СН2)4(СН=СНСН2)3СН=СН(СН2)3СООН |
Жирные кислоты арахидоновая (С20:4), линолевая (С18:2) и линоленовая (С18:3) называют незаменимыми, поскольку они не синтезируются в клетках млекопитающих и должны поступать туда с пищей. Вокруг каждой двойной С=С связи возможны две стереоизомерные конфигурации (рисунок 55(б, в), рисунок 56(6)). Цис-двойная связь обеспечивает жесткий излом (кинк) в, вообще говоря, гибком углеводородном "хвосте» жирной кислоты.
Основная функция триацилглицеролов в живых организмах - это запасание энергии. Для этого они накапливаются в цитозоле клеток запасающих тканей (например, подкожной клетчатки) в виде мелкодисперсной эмульсии масляных капелек и могут занимать почти весь объем запасающей клетки.
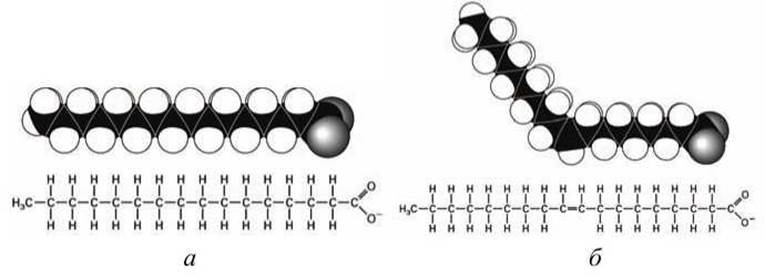
Рисунок 56 - Влияние двойной С=С связи на форму жирной кислоты: а - пальмитат (ионизированная форма пальмитиновой кислоты); б - олеат (ионизированная форма олеиновой кислоты)
В расчете на единицу веса триацилглицеролы запасают вдвое больше энергии, чем углеводы, и, накапливаясь в больших количествах, могут обеспечивать энергетический обмен организма в течение нескольких недель, тогда как в форме гликогена организм может запасти энергию не более чем на сутки.
Однако энергия, запасенная в углеводах, становится доступна для использования организмом быстрее, чем запасенная в форме триацилглицеролов. Поэтому полисахариды (крахмал и гликоген) и жиры (триацилглицеролы) работают как оперативная и долговременная системы запасания энергии соответственно.
Дополнительно триацилглицеролы могут служить для теплоизоляции и как источник воды: при окислении триацилглицеролов образуется вдвое больше воды, чем из углеводов, и это используется пустынными животными.
Толстый подкожный слой жировой клетчатки надежно защищает тюленей, моржей, пингвинов и других полярных теплокровных животных от холода.
Фосфолипиды - сложные липиды, похожие на триацилглицеролы наличием жирных кислот, связанных с глицеролом. Однако в фосфолипидах глицерол связан не с тремя, а лишь с двумя цепями жирных кислот, а оставшееся свободное место в молекуле глицерола занимает фосфатная группа, которая в свою очередь соединена с молекулой полярного спирта, различного для разных фосфолипидов. Полярный спирт образует как бы "голову" молекулы фосфолипида, к которой через глицерол присоединены два неполярных "хвоста" (рисунок 57)
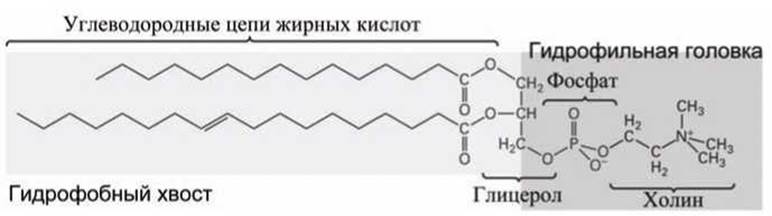
Рисунок 57 - Фосфатидилхолин - типичный фосфолипид
Примечательно, что самые большие пространственные структуры, которые формируются в клетке, образованы не из макромолекул, таких, как белки или нуклеиновые кислоты, а агрегацией липидов в биомембраны - обособленную жидкую фазу. Липиды, которые используются клеткой, устроены так, чтобы осуществлялась самопроизвольная агрегация, в результате которой в клетке формируется клеточная инфраструктура.
В молекуле липидов объединены два противоположных химических качества. Они одновременно содержат и полярную или даже заряженную гидрофильную группу атомов, которая стремится раствориться в воде, и одну или несколько гидрофобных углеводородных цепей, которые выталкиваются из воды (рисунок 57). Такие молекулы называются амфифильными.
Двойственная природа липидных молекул приводит к их самоорганизации в мембранные структуры, в которых заряженные (или полярные) головки обращены в сторону воды, а углеводородные хвосты упакованы внутри мембраны (рисунок 58).
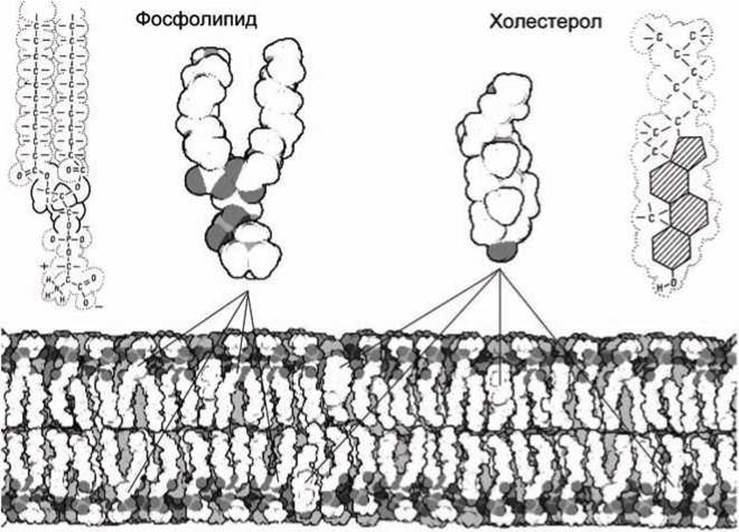
Рисунок 58 - Молекулы фосфолипидов (фосфатидилсерин) и холестерола из которых происходит самосборка биомембраны. Темно-серым цветом обозначены полярные участки молекул
При этом основными межмолекулярными взаимодействиями, обеспечивающими самосборку липидов в биомембрану, являются неспецифические нековалентные взаимодействия: ван-дер-ваальсовые, электростатические, водородные и гидрофобные.
Наиболее распространенные типы липидов в природе это фосфолипиды и гликолипиды. В основе их конструкции лежит молекула глицерола, имеющая три гидроксильные группы, вместо которых могут быть присоединены три другие группы. Две из них, как правило, это жирные кислоты, присоединенные к глицеролу через карбоксильную группу. Вместо третьей гидроксильной группы к глицеролу присоединяется фосфатная группа или другая полярная (или заряженная) группа. Если жирнокислотные хвосты имеют в своем составе ненасыщенные углерод-углеродные связи, то в таких местах образуются жесткие кинки (рисунок 56(6)).
Жирнокислотные хвосты с кинками гораздо хуже упаковываются в упорядоченную структуру, и, поэтому, биомембрана с ненасыщенными углеводородными цепями в липидах имеет более низкую температуру фазового перехода из физиологического жидкокристаллического в замороженное гель-состояние, чем у биомембраны с насыщенными С-С связями в липидах.
Холестерол и другие стеролы устроены иначе. Они состоят из нескольких жестко связанных гидрофобных углеводородных колец протяженностью такой же, как и углеводородные хвосты фосфолипидов. Гидроксил на одном из концов обеспечивает гидрофильность, ориентируя холестерол в мембране. Холестерол добавляется к мембранам в разных пропорциях для того, чтобы модифицировать свойства биомембран. Поскольку молекула холестерола является жесткой, то добавление холестерола ингибирует (затрудняет) движение соседних липидов, увеличивая тем самым вязкость мембраны и делая её менее проницаемой для малых молекул.
Липиды используются клетками для формирования мембранных структур клетки.
Биомембраны непроницаемы для ионов и больших молекул, начиная с сахаров и полипептидов, но легко проницаемы для молекул воды, кислорода, азота. Примечательно, что молекулы, содержащие много атомов углерода, достаточно легко проникают через мембраны. Именно поэтому алкоголь легко распространяется по организму, пересекая все барьеры.
При взбалтывании в воде или водных растворах молекулы липидов спонтанно формируют мицеллы, в которых углеводородные неполярные хвосты липидов спрятаны от воды, а полярные головы располагаются на поверхности частицы, взаимодействуя с водным окружением (рисунок 59(a)).
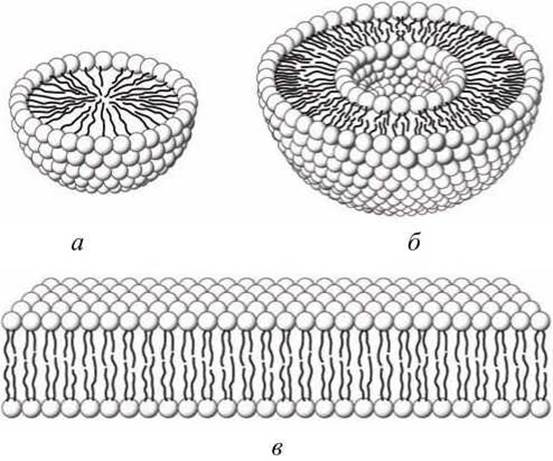
Рисунок 59 - Поперечное сечение трех структур, формируемых липидами в водных растворах: а - мицелла; б - липосома; в - плоская липидная мембрана
На поверхности раздела двух водных фаз липиды самопроизвольно формируют бислои.
В бислоях углеводородные "хвосты" направлены внутрь от водной фазы и образуют непрерывный гидрофобный углеводородный слой, а располагающиеся снаружи гидрофильные "головы" погружены в водный раствор (рисунок 59(b)).
При сильном встряхивании бислоев из них формируются липосомы - замкнутые пузырьки, окруженные непрерывным липидным бислоем (рисунок 59(6)).
Плотная упаковка молекул липидов в биомембранах обеспечивается сочетанием гидрофобных сил со стороны водного окружения и ван-дер-ваальсовых взаимодействий между молекулами липидов. Липидный бислой является основным структурным компонентом всех биомембран.
Хотя в мембранах присутствуют и другие молекулы: холестерин, гликолипиды, белки, - именно гидрофобный углеводный слой, формируемый липидными молекулами, обеспечивает главную, барьерную, функцию биомембран, которая, в свою очередь, обеспечивает компартментацию клеточных субструктур.
Вопросы для самоконтроля
1. Запишите общую формулу углеводов.
2. В чем сходство и различие рибозы и дезоксирибозы?
3. В чем сходство и различие а- и ß-гликозидной связей?
4. В чем сходство и различие гликогена и целлюлозы?
5. В чем сходство и различие гликогена и крахмала?
6. Что такое гликопротеины и чем они отличаются от протеогликанов?
7. В чем сходство и различие гликогена и триацилглицеролов?
8. Перечислите основные жирные кислоты фосфолипидов?
9. В чем сходство и различие фосфолипидов и триацилглицеролов?
10. Какие межмолекулярные взаимодействия обеспечивают формирование биомембран?