Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Компоненты биомолекулярных комплексов
Углеводы
На долю углеводов приходится до 80% сухого вещества некоторых растительных тканей и до 20% некоторых животных тканей. Простейшими углеводами, встречающимися в живых организмах, являются моносахариды, имеющие общую формулу Сn(Н2O)n, где n = 3-7 (именно этим объясняется их общее название "углеводы"). Практически все природные моносахариды, так же как и аминокислоты, могут существовать в D- форме и в L-форме (рисунок 46). В отличие от аминокислот, почти все моносахариды живых организмов относятся к D-ряду.
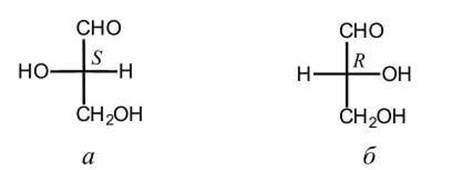
Рисунок 46 - Энантиомеры глицеринового альдегида СН2(ОН)СН(ОН)СНО: а - L-глицериновый альдегид; б - D-глицериновый альдегид
Моносахариды, содержащие пять и более атомов углерода, в растворах существуют как в линейной форме, так и в виде замкнутых циклических структур (рисунок 47).
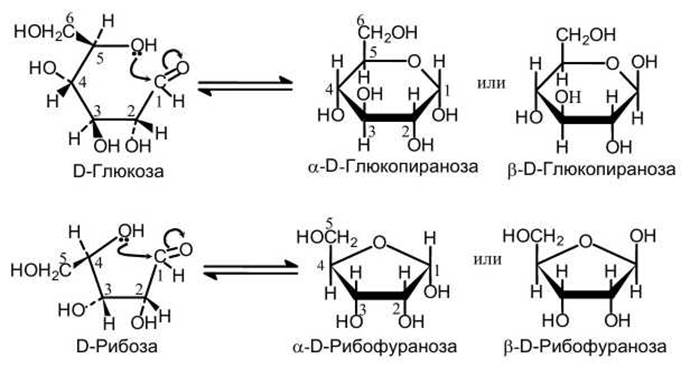
Рисунок 47 - Образование циклической формы моносахаридов
При этом замкнутые структуры могут существовать в виде двух стереоизомеров a и ß (рисунок 47).
В цитозоле молекула моносахарида постоянно "переключается" между линейной формой и двумя циклическими стереоизомерами. Например, моносахарид глюкоза переключается между циклическими изомерами а-глюкоза и ß-глюкоза через линейный изомер (рисунок 48).
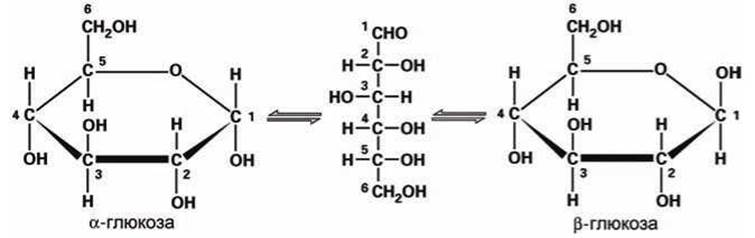
Рисунок 48 - Переключение изомеров глюкозы
Следует иметь в виду, что в действительности шести- и пятичленные кольца неплоские. Шестичленное кольцо большинства сахаров имеет конфигурацию кресла (рисунок 49). Неплоской является и фуранозная форма.
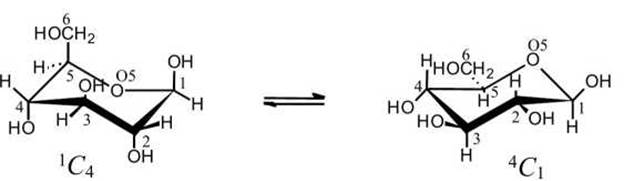
Рисунок 49 - Две конформации кресла ß -D-глюкопиранозы. Буква "С" означает конформации кресла (Chair); цифры указывают, какой из атомов отогнут вверх (надстрочный индекс), а какой - вниз (подстрочный индекс) при каноническом написании проекции
Обычно в равновесной смеси преобладает тот конформер, в котором наибольшее число объемистых заместителей располагаются на экваториальных связях. ß-D-Глюкопираноза существует в растворе практически полностью в виде 4Сгконформера (все объемистые заместители расположены на экваториальных связях).
Основными моносахаридами живых организмов являются глюкоза, фруктоза, рибоза (входит в состав РНК) и 2-дезоксирибоза (входит в состав ДНК), а также галактоза, манноза и рибулоза (рисунок 50).
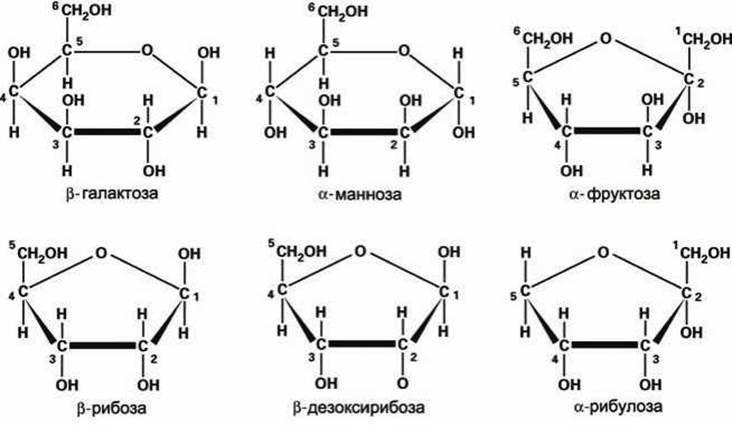
Рисунок 50 - Схемы основных моносахаридов
Моносахариды могут соединяться друг с другом, образуя гликозидную связь, которая образуется между гидроксильной группой одного моносахарида и альдегидной группой или кетогруппой другого моносахарида. При этом отщепляется молекула воды, и образуется дисахарид (рисунок 51).
Присоединение аналогичным путем дополнительных моносахаридов приводит к образованию олигосахаридов всё возрастающей длины (трисахаридов, тетрасахаридов и т.д.) вплоть до очень больших молекул полисахаридов, содержащих сотни и тысячи моносахаридных остатков.
Полисахариды, которые называют также гликанами, отличаются друг от друга как природой составляющих их моносахаридных остатков, так и длиной и степенью разветвленности цепей.
Поскольку у каждого моносахарида имеется несколько свободных гидроксильных групп, способных образовывать связь с другим моносахаридом или каким-либо иным соединением, число возможных структур полисахаридов исключительно велико.
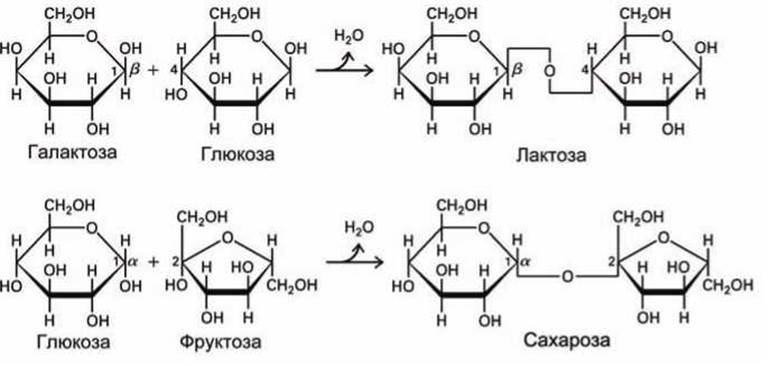
Рисунок 51 - Схема образования дисахаридов
Поскольку каждая из гидроксильных групп может находиться с разных сторон от плоскости кольца (а- и ß-положения), то каждую конкретную гликозидную связь обозначают как а/β-m, n-связь, где m и n обозначают номер углеродного атома, несущего гидроксильную группу, участвующую в образовании гликозидной связи. Например, на рисунке 51 показаны ß-1,4- и а-1,2-гликозидные связи.
Даже простейший дисахарид из двух остатков глюкозы может быть образован одиннадцатью различными гликозидными связями, а три различные гексозы (например, глюкоза, фруктоза и галактоза), соединяясь между собой, способны образовать несколько тысяч различных трисахаридов.
Структурная функция полисахаридов. В живых организмах углеводы выполняют структурную, энергетическую и специальные функции. Основными структурными полисахаридами служат: у растений - целлюлоза и пектины, а у животных и грибов — хитин.
Целлюлоза - самое распространенное органическое соединение на Земле, поскольку из нее построены клеточные стенки растений. В частности, древесина и хлопок почти целиком состоят из целлюлозы. В день на каждого живущего на Земле человека растения синтезируют примерно 50 кг целлюлозы.
Целлюлоза - прочное, волокнистое, водонерастворимое вещество, каждая молекула которого состоит из 104 и более остатков D-глюкозы, соединенных в неразветвленную цепь посредством β-1,4-гликозидных связей (рисунок 52). Полимерные цепи целлюлозы сильно вытянуты и соединены друг с другом водородными связями.
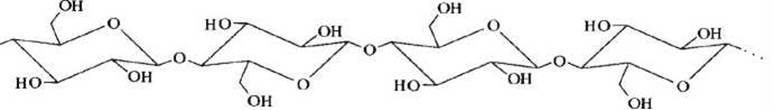
Рисунок 52 - Схема полисахаридной цепи целлюлозы
Хитин, как и целлюлоза, - линейный неразветвленный полисахарид, однако структурными единицами его служит не D-глюкоза, а N-ацетил-D-глюкозамин (рисунок 53).
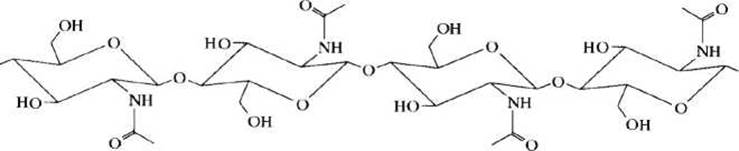
Рисунок 53 - Схема полисахаридной цепи хитина
Из хитина построены прочные нерастворимые покровы ракообразных и насекомых, а также клеточные стенки грибов. Хитиновый каркас многих ракообразных усилен за счет включений карбоната кальция.
Энергетическая функция полисахаридов. Углеводы служат главным оперативным источником энергии в клетках. В результате окисления глюкоза и другие моносахариды распадаются до СО2 и Н2O, и высвобождающаяся при этом химическая энергия используется клеткой.
Гликолиз — это практически универсальный процесс, в котором шестиуглеродная молекула глюкозы (С6Н12O6) в результате серии ферментативных превращений в цитозоле анаэробно превращаются в две трёхуглеродные молекулы пирувата (С3Н5О3). При этом расходуются две молекулы АТФ, но четыре молекулы АТФ синтезируются в ходе гликолиза, поэтому гликолиз одной молекулы глюкозы сопровождается синтезом двух молекул АТФ. Кроме того, восстанавливаются две молекулы никотинамид адениндинуклеотида НАД:
Глюкоза + 2НАД+ +2АДФ + 2Р → 2 пирувата + 2АТФ + 2НАДН + Н+.
Для запасания энергии впрок используются полисахариды, построенные из повторяющихся остатков глюкозы, - крахмал (у растений) и гликоген (у животных).
Когда необходима энергия, молекулы глюкозы отщепляются от крахмала или гликогена, а при избытке глюкозы её молекулы присоединяются к полимерным цепям крахмала или гликогена и удлиняют их. Таким образом, резервные полисахариды всё время меняют свой размер в зависимости от потребности организма в энергии.
Крахмал представляет собой смесь двух полимеров D-глюкозы — а-амилозы и амилопектина (рисунок 54).
а-Амилоза состоит из длинных неразветвленных цепей с молекулярной массой от нескольких тысяч до 5x105, построенных из остатков D-глюкозы, соединенных а-1,4-гликозидными связями.
Амилопектин также имеет высокую молекулярную массу, но, в отличие от а-амилозы, его молекулы сильно разветвлены. В неразветвленных участках амилопектина остатки D-глюкозы соединены а-1,4-гликозидными связями, a в местах ветвления - а-1,6-гликозидными связями.
Гликоген - сильно разветвленный полисахарид, так же, как и амилопектин, состоящий из остатков D-глюкозы, связанных а-1,4-гликозидными связями, a в местах ветвления - а-1,6-гликозидными связями.
Больше всего гликогена содержится в клетках печени, где на его долю приходится до 7% общего веса органа. В клетках печени гликоген присутствует в виде крупных гранул, состоящих в свою очередь из меньших гранул, каждая из которых образована одной сильно разветвленной молекулой со средней молекулярной массой в несколько миллионов. С этими гранулами прочно связаны ферменты, ответственные за синтез и распад гликогена.
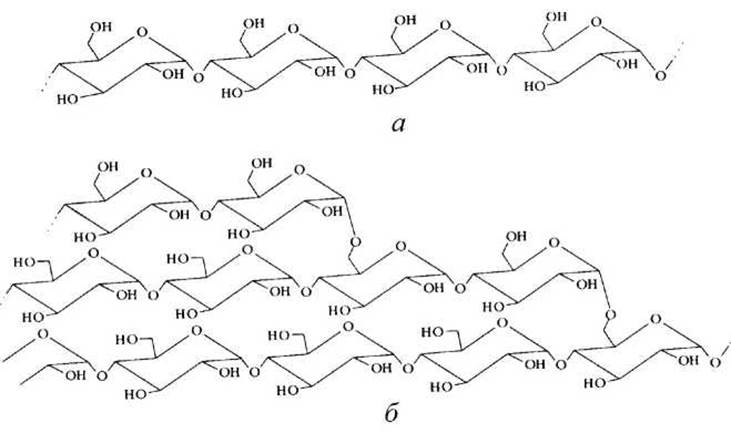
Рисунок 54 - Схема полисахаридов крахмала: а - амилоза; б - амилопектин
Хотя и целлюлоза, и крахмал, и гликоген состоят из остатков глюкозы, они сильно различаются по своим свойствам из-за различия гликозидных связей, которыми соединены остатки глюкозы в этих молекулах.
Благодаря геометрическим особенностям а-1,4-гликозидных связей линейные участки полимерных цепей в молекулах гликогена и крахмала стремятся принять скрученную, спиральную конформацию, что способствует образованию плотных гранул крахмала и гликогена, обнаруживаемых в клетках.
Напротив, из-за структурных особенностей ß-1,4-гликозидпых связей молекулы целлюлозы имеют сильно вытянутую линейную конформацию.
а-1,4-связи легко гидролизуются ферментом амилазой, поэтому крахмал легко усваивается животными и человеком, расщепляясь до D-глюкозы. Однако в кишечнике животных нет фермента, способного гидролизовать β-1,4-гликозидные связи, поэтому целлюлоза не переваривается, и её D-глюкозные остатки не могут служить пищей для большинства организмов.
Тем не менее, жвачные и термиты используют целлюлозу в качестве пищи благодаря обитающим в их кишечнике микроорганизмам (бактериям и простейшим), которые синтезируют целлюлазу - фермент, гидролизующий ß-1,4-гликозидпые связи. Целлюлазу синтезируют также некоторые грибы, вызывающие гниение древесины.
Специальные функции полисахаридов. К полисахаридам со специальными функциями относятся очень сложные соединения, биохимические функции которых не всегда известны, например, камеди и слизи.
Углеводы могут ковалентно связываться с белками и липидами, образуя гликопротеины и гликолипиды.
Как правило, такие гибридные молекулы входят в состав оболочек клеток, и их олигосахаридная часть участвует в процессах межклеточного узнавания и рецепции сигнальных молекул.
Жесткость олигосахаридных структур, их растворимость в воде и огромное разнообразие возможных конфигураций делают их наиболее подходящими для выполнения этих функций.
Протеогликанами (или пептидогликанами) называются гликопротеины, в которых на долю полисахарида приходится основная часть молекулы - более 95%. Они входят в состав основного вещества, заполняющего пространство между клетками в большинстве тканей, а у бактерий (прокариоты) именно пептидогликаны (но не хитин и целлюлоза) служат структурной основой клеточных стенок. В эукариотической клетке клеточная стенка, если она есть может содержать хитин или целлюлозу (но не пептидогликан).
Большое количество гидроксильных групп в полисахаридах образуют водородные связи с другими донорами или акцепторами, обеспечивая существование двух важных типов структур в живых организмах.
В одном случае, отдельные полисахаридные цепи ассоциируются с большим количеством молекул воды, образуя клейкий гель. В таком виде углеводы покрывают большинство наших клеток, образуя их клейкую защитную оболочку. Гликопротеины слизи дают представление о внешнем виде и свойствах такого образования.
В другом случае, углеводные цепи прочно ассоциируются друг с другом посредством сети водородных связей, образуя прочные волокна, внутри которых практически нет воды. В таком виде полисахариды используются для формирования макроскопической инфраструктуры клеток и межклеточных образований и для запасания энергии. Некоторые из наиболее впечатляющих биологических структур, созданных природой, включая прочные стволы гигантских деревьев и жесткие водонепроницаемые панцири членистоногих, обязаны своей силой и прочностью полисахаридам.