ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
11. ОСНОВНІ МЕХАНІЗМИ ОБМІНУ РЕЧОВИН І ПЕРЕТВОРЕННЯ ЕНЕРГІЇ У МІКРООРГАНІЗМІВ
11.6. ДИХАЛЬНИМ ЛАНЦЮГ І ФОСФОРИЛЮВАННЯ (СИНТЕЗ АТФ) ПРИ ПЕРЕНЕСЕННІ ЕЛЕКТРОНІВ
11.6.5. Механізм синтезу АТФ при перенесенні електронів
Як синтезується АТФ при перенесенні електронів у дихальному ланцюгу? Звідки береться енергія, яка потім акумулюється в макроергічних зв'язках АТФ? Яким є механізм синтезу АТФ при перенесенні електронів? Відповідь на ці запитання дає хіміоосмотична гіпотеза, запропонована англійським ученим П. Мітчелом у 1961 р. Основні її положеннями такі (рис. 11.11):
мембрана, в якій локалізований дихальний ланцюг, є непроникною для протонів та гідроксильних іонів;
віддані субстратами відновлювальні еквіваленти переносяться на плазматичну мембрану чи на внутрішню мембрану мітохондрій. Взаєморозміщення компонентів дихального ланцюга у мембрані є таким, що при транспорті електронів віл субстрату до кисню протони зв’язуються всередині мембрани, а вивільнюються назовні. Можна уявити, що електрони у мембрані проходять зигзагоподібний шлях і при цьому переносять протони зсередини назовні. Так, окиснення НАДН у дихальному ланцюгу супроводжується виділенням протонів на зовнішньому боці мембрани (рис. 11.11,а)
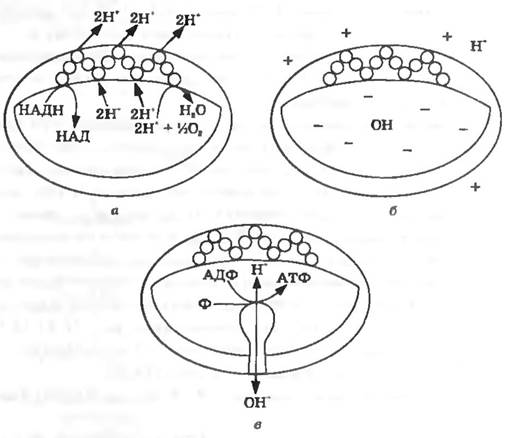
Рис. 11.11. Хіміо-осмотична гіпотеза окиснювального фосфорилювання в мітохондріях
наслідком такої транслокаціі протонів є встановлення на мембрані градієнта pH (ΔρΗ) та градієнта електричного заряду (Δψ) з позитивним потенціалом назовні та негативним всередині (рис. 11.11,б). Отже, внутрішній простір мітохондрій або внутрішній бік плазматичної мембрани у бактерій є електронегативними по відношенню до зовнішнього середовища і відзначаються більш високим pH. Трансмембранна різниця електричного заряду (електричного потенціалу) та хімічного градієнта концентрації іонів водню (градієнт pH) створюють трансмембранний електрохімічний градієнт протонів (протонрушійна сила, протонний потенціал). Протонрушійна сила може бути зумовлена або тільки градієнтом pH, або тільки градієнтом електричного заряду;
для розрядки потенціалу, що виникає в певних місцях плазматичної мембрани (чи внутрішньої мембрани мітохондрій), вбудований Н' -залежний АТФ-синтазний ферментний комплекс, який каталізує реакції синтезу чи гідролізу АТФ (рис. 11.11, в). Реакція синтезу АТФ спряжена з транспортом протонів за градієнтом протонного потенціалу (з зовнішнього боку мембрани на внутрішній), в результаті чого відбувається його розрядка. Енергія, яка виникає при цьому (енергія, яка генерується протонрушійною силою) і акумулюється у макроергічних зв'язках АТФ.
Реакція гідролізу АТФ супроводжується перенесенням протонів проти градієнта, в результаті чого зовні мембрани накопичуються протони і утворюється або збільшується ΔμΗ. Відбувається взаємне перетворення двох форм енергії: ΔμΗ → АТФ. Отже, можна сказати, що АТФ-синтазний комплекс відіграє роль «протонного насоса».
Слід зазначити, що енергія протон рушійної сили використовується не тільки для синтезу АТФ. Ця форма енергії забезпечує багато які процеси, локалізовані на мембрані у бактерій: активний транспорт, рух джгутиків, зворотне перенесення електронів та ін. Для деяких процесів ця форма енергії зручніша, ніж АТФ, тому що енергія протонрушійної сили не міститься у вигляді певних порцій, як в АТФ, тому не існує нижньої межі для її утворення. Вона може утворюватись і споживатись в умовах, коли синтез АТФ є неможливим.
Отже, у мікроорганізмів існує дві форми енергії: енергія у вигляді АТФ та енергія протонрушійної сили. АТФ утворюється при субстратному фосфорилюванні та фосфор плюванні при перенесенні електронів. Енергія протонрушійної сили генерується під час транспорту електронів у дихальному ланцюгу або у процесі гідролізу АТФ за участю АТФ-синтазного ферментного комплексу.