ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
20. РЕГУЛЯЦІЯ МЕТАБОЛІЗМУ
20.3. МЕХАНІЗМИ СИНТЕЗУ ФЕРМЕНТІВ (ІНДУКЦІЇ ТА РЕПРЕСІЇ)
20.3.1. Індукція лактозного оперону
Найдетальніше механізми індукції та репресії досліджені у процесі синтезу ферментів катаболізму лактози у Е. coli (рис. 20.6).
Три гени, які кодують β-гaлактозидозу, пермеазу та трансацетилазу (структурні гени) локалізовані у хромосомі поряд один з одним. Ділянки ДНК. до яких приєднаються регуляторні білки, безпосередньо прилягають до структурних генів. Ці області називаються промотором (Р) та оператором (О). Промотор являє собою послідовність основ, яка розпізнається ДНК залежною РНК полімеразою. Промотор є місцем зв’язування РНК-полімерази, з нього починається транскрипція. Оператор являє собою нуклеотидну послідовність, яка розміщується між промотором і структурними генами. Оператор взаємодіє з регуляторним білком-репресором, від якого залежить, чи буде подавлена транскрипція, чи ні. Промотор, оператор і структурні гени утворюють оперон. Опероном наливають групу функціонально зв’язаних між собою генів.
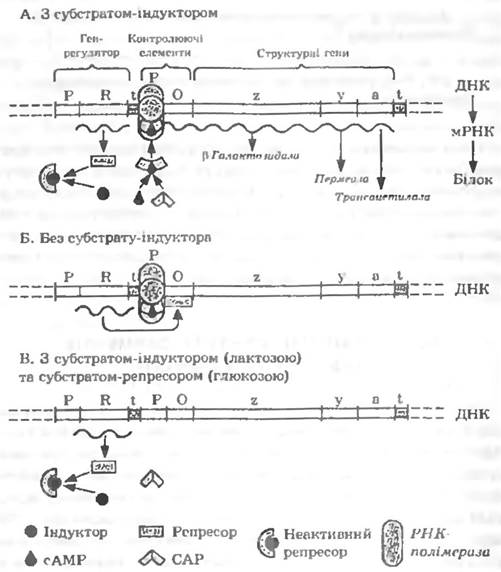
Рис 20.6. Модель регуляції лактозного оперону
За синтез регуляторних білків відповідають регуляторні гени (Р та R; на рис. 20.6 розміщені ліворуч від промоторе). Вони не обов’язково можуть розміщуватись поряд з відповідним опероном. Гени-регулятори, як правило, є конститутивними. За припинення (термінацію) синтезу мРНК відповідальним є термінатор (t) — спеціальна область ДНК, яка розміщується біля кінця оперону. Вся описана структура називається лактозним опероном. Розглянемо, як функціонує лактозний оперон. На рис. 20.6 наведено три можливі варіанти: без індуктора (лактози); з індуктором (лактозою); катаболітна репресія (при сутні два субстрати — лактоза та глюкоза).
У першому випадку (рис. 20.6, Б) (за відсутності лактози) репресор зв’язується з оператором і перешкоджає транскрипції, тобто операторна область блокується рспресором.
Коли в клітину потрапляє лактоза, з неї за участю β-галактозидази утворюється алолактоза, яка служить внутрішнім індуктором. Цей внутрішній індуктор зв’язується з репресором, внаслідок чого репресор піддається конформаційним змінам так, що не може зв’язуватись з оператором (рис. 20.6, А). У результаті цього оперон залишається вільним, транскрипція стає можливою. Це так звана негативна регуляція. За негативної регуляції для ініціювання транскрипції репресор повинен бути витіснений індуктором.
Існує і позитивна регуляція, за якої необхідною умовою транскрипції є взаємодія оперона з активатором — другим регуляторним білком CAP або CRP (puc. 20.6, A). CAP (catabolitc activator protein) i CRP (cyclic AMP receptor protein) є синонімами. Транскрипція можлива тільки тоді, коли білок CAP зв'язаний з промотором. Таке зв’язування CAP з промотором є необхідною умовою для приєднання РНК-полімерази до ДНК. Проте в свою чергу CAP може зв’язуватись з промотором тільки у тому разі, якщо в клітині в достатньо високій концентрації присутній циклічний АМФ (сАМР).
У третьому випадку (катаболітна репресія лактозного оперона; рис. 20.6, В), коли у середовищі присутні лактоза та глюкоза, синтез ферментів лактозного оперону пригнічується. Така дія глюкози зумовлена тим. що в її присутності концентрація циклічного АМФ є дуже низькою. У таких умовах CAP не може зв’язуватись з промотором, не відбувається приєднання РНК полімерази до ДНК і процесу транскрипції. Відповідно до цього механізму катаболітна репресія синтезу ферментів лактозного оперону у присутності глюкози може бути знята, якщо добавити у середовище циклічний АМФ у високих концентраціях.