Общая микробиология - Шлегель Г. 1987
Грибы
Своим названием вся эта группа организмов обязана самым заметным ее представителям - шляпочным грибам (греч. mykes, лат. fungus). Грибы относятся к эукариотам. С растениями их сближает ряд общих признаков: наличие клеточной стенки и вакуолей, заполненных клеточным соком; хорошо видимое под микроскопом движение протоплазмы; неспособность к активному перемещению. У грибов, однако, нет фотосинтетических пигментов; это С-гетеротрофы (точнее, хемоорганогетеротрофы). Грибы растут в аэробных условиях и получают энергию путем окисления органических веществ. По сравнению с растениями, имеющими стебель, корни и листья, грибы слабо дифференцированы морфологически, и у них почти нет разделения функций между разными частями организма.
Вегетативное тело. Вегетативное тело (таллом) гриба состоит из нитей толщиной около 5 мкм, сильно разветвленных и разрастающихся по поверхности или во всем объеме питательного субстрата. Эти так называемые гифы состоят из клеточных стенок и цитоплазмы с ее включениями. Гифы либо не имеют поперечных перегородок (у низших грибов), либо разделены такими перегородками (септами) на клетки (у высших грибов). В последнем случае, однако, цитоплазма одной клетки сообщается с цитоплазмой соседней клетки через пору, находящуюся в центре перегородки (рис. 5.1).
Всю совокупность гиф грибного таллома называют мицелием. На определенных стадиях, например при переходе в фазу бесполого или полового размножения, мицелий образует плотные, похожие на паренхиматозную ткань сплетения, так называемую плектенхиму. Типичный пример плектенхимы - мясистые плодовые тела шляпочных грибов. У высших грибов мицелий образует также плотные тяжи - ризоморфы, функция которых связана с транспортом веществ.
Рост и размножение. Гифы грибов растут путем удлинения кончиков (апикальный рост). У большинства грибов любая часть мицелия способна к росту. Для посева достаточно маленького кусочка мицелия - из него образуется новый таллом. Структуры и механизмы, обеспечивающие размножение, исключительно многообразны и служат основой для классификации грибов. Различают два типа размножения - половое и бесполое. Большинство грибов размножается обоими способами.
Бесполое размножение осуществляется обычно при помощи спор, путем почкования или фрагментации. Наиболее широко распространено и наиболее дифференцировано спорообразование. На концах гиф отшнуровываются конидиоспоры (у Penicillium, Aspergillus). Если споры образуются внутри спорангиев, то говорят о спораигиоспорах (у Мuсоr, Rhizopus). У низших грибов спорангиоспоры часто движутся с помощью жгутиков; в таком случае их называют зооспорами. Строение жгутиков такое же, как у всех эукариот: они отходят от лежащих в цитоплазме блефаропластов и состоят из 11 параллельных фибрилл, из которых 9 периферийных фибрилл расположены вокруг двух, находящихся в центре (структура типа 9 + 2).
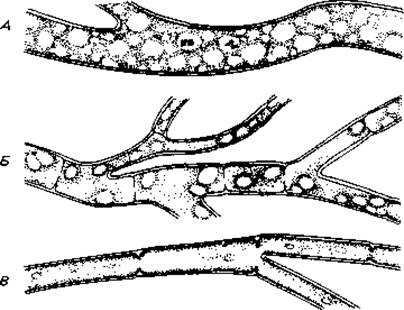
Рис. 5.1. Вегетативные гифы грибов. А. Гифы фикомицетов несептированные, т. е. не имеют поперечных перегородок. Б. Для эумицетов характерны гифы с перегородками - септированные. В. У оомицета Leptomitus lacteus гифы разделены неполными перетяжками.
Для дрожжей характерным видом бесполого размножения является почкование. При этом на материнской клетке образуется небольшая выпуклость - почка, в которую переходит одно ядро, после чего почка отшнуровывается (рис. 5.2). Размножение может осуществляться также путем разлома гиф на отдельные клетки - оидии или артроспоры (как, например, у молочной плесени Endomyces lactis). У некоторых грибов такие клетки окружаются толстой стенкой, и в этом случае их называют хламидоспорами. Наконец, некоторые представители дрожжей (Schizosaccharomyces), подобно бактериям, размножаются делением надвое.
Половое размножение у грибов, так же как у других эукариот, включает слияние двух ядер. Такое слияние ядер у разных грибов происходит через различные промежутки времени после первого контакта между родительскими клетками. В процессе полового размножения можно различить три фазы. Прежде всего происходит плазмогамия - соединение двух протопластов. Возникшая в результате этого клетка содержит два ядра. Эта пара ядер (дикарион) не обязательно сливается сразу же. Во время последующих делений клетки могут оставаться в дикариотической фазе. Оба ядра делятся при этом одновременно (сопряженное деление). Лишь позднее, часто только после образования плодового тела, происходит слияние обоих гаплоидных ядер (кариогамия) с образованием диплоидного ядра зиготы. За кариогамией следует мейоз, или редукционное деление, при котором число хромосом уменьшается до исходного (гаплоидного). Три названных процесса - плазмогамия, кариогамия и мейоз - у некоторых грибов следуют непосредственно один за другим, у других же происходят на самых различных стадиях развития гриба.
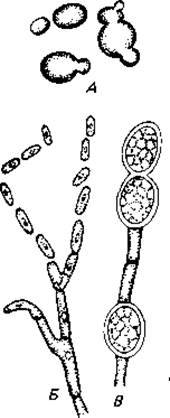
Рис. 5.2. Бесполое размножение некоторых грибов. А. Почкование (у дрожжей). Б. Разделение гиф на отдельные клетки, называемые оидиями или артроспорами и ведущие себя как споры (у Collybia sp.). В. Образование толстостенных хламидоспор (у Fusarium). (Alexopoulos С. J., Einführung in die Mykologie, Stuttgart: Fischer, 1966.
У низших грибов фаза полового размножения начинается с образования половых клеток, или гамет. Если гаметы, происходящие от мужской и женской родительских клеток, морфологически неразличимы, их называют изогаметами. Гаметы образуются часто в особых морфологически дифференцированных клетках - гаметангиях. Если эти последние различны по своей форме, то мужские гаметангии называют антеридиями, а женские - оогониями.
По способу переноса гамет и осуществления плазмогамии различают несколько типов грибов. У низших, преимущественно водных, грибов обе гаметы подвижны (планогаметы) и слияние их происходит вне гаметангиев. У оомицетов подвижна только мужская гамета; она проникает в оогоний и оплодотворяет яйцеклетку. Для зигомицетов характерна гаметангиогамия слияние целых соприкасающихся друг с другом многоядерных гаметангиев в многоядерную ценозиготу.
Если женские и мужские гаметангии образуются на одном и том же вегетативном теле, развившемся из одной споры, то говорят о гомоталлических (гермафродитных) грибах. У гетероталлических грибов талломы различны в половом отношении, т. е. несут либо только мужские, либо только женские половые органы. У гомоталлических грибов возможно самооплодотворение (аутогамия). В тех случаях, когда самооплодотворению препятствует какого-то рода физиологический барьер, говорят об их несовместимости. Такая несовместимость наблюдается, например, у Neurospora. Хотя у этого гриба на одном и том же мицелии образуются гаметангии обоего пола, оплодотворение может происходить только между мицелиями разных половых знаков (+ и —). Мицелии одного и того же знака несовместимы.
Классификация. Классификация грибов, так же как и бактерий, преследует в основном практические цели; однако она учитывает при этом и филогенетические связи. Номенклатура бинарная: каждому виду присваивается родовое и видовое название (например, Aspergillus niger). Виды объединяются в роды, роды - в семейства (-асеае), семейства - в порядки (-ales), порядки - в классы (-mycetes). К отделу Mycota, или Fungi, т. е. к грибам, относятся истинные слизевики (миксомицеты), низшие грибы (фикомицеты) и, наконец, высшие грибы (собственно грибы, или эумицеты).
Полный обзор таксономии, морфологии и физиологии грибов читатель может найти в ряде руководств (Мюллера и Леффлера, Геймана, Алексопулоса, фон Аркса и др.). К этим руководствам и следует обращаться микологам и микробиологам, особо интересующимся отдельными группами грибов. Здесь мы можем рассмотреть лишь немногих представителей различных групп - формы, которые либо служат модельными системами при разного рода исследованиях, либо имеют большое практическое значение (табл. 5.1).