Общая микробиология - Шлегель Г. 1987
Основные механизмы обмена веществ и преобразования энергии
Поглощение веществ клетками
Для того чтобы экзогенный субстрат мог быть использован клеткой, он должен пройти через ее пограничные слои. Клеточная стенка не служит существенной преградой для небольших молекул и ионов, но она задерживает макромолекулы, масса которых превышает 600 Да. Пограничным слоем, ответственным за транспорт питательных веществ внутрь клетки, является плазматическая мембрана.
Перенос питательных веществ через плазматическую мембрану, как правило, специфичен: поглощаться могут только те вещества, для которых имеется соответствующая транспортная система. За небольшими исключениями, транспорт зависит от наличия специфических пермеаз или транслоказ. Речь идет о мембранных белках, само название которых указывает на то, что они обладают свойствами ферментов, т. е. могут индуцироваться субстратом, специфичны в отношении субстрата и образуются только в таких условиях, в которых возможен синтез белков.
Что касается механизма транспорта веществ, то различают ряд различных процессов, два из которых способны обеспечивать только транспорт, но не накопление веществ в клетке; им можно противопоставить процессы активного транспорта, приводящие к аккумуляции веществ внутри клетки (рис. 7.18 и 7.19).
Простая диффузия. Неспецифическое проникновение веществ в клетку происходит путем пассивной диффузии. Для диффузии существенны величина молекул и степень их липофильности. Скорость перемещения путем диффузии невелика. Для сахаров такие процессы не были обнаружены, и они мало вероятны. Путем простой диффузии в клетку проникают, видимо, яды, ингибиторы и другие чуждые клетке вещества.
Облегченная диффузия. При облегченной диффузии вещество, содержащееся в питательной среде, транспортируется в клетку «вниз» по своему градиенту концентрации. Этот процесс осуществляется благодаря субстрат-специфической пермеазе и не требует затраты метаболической энергии. Скорость транспорта в широком диапазоне зависит от концентрации субстрата в среде (рис. 7.19). Питательное вещество не может накапливаться в клетке против градиента концентрации.
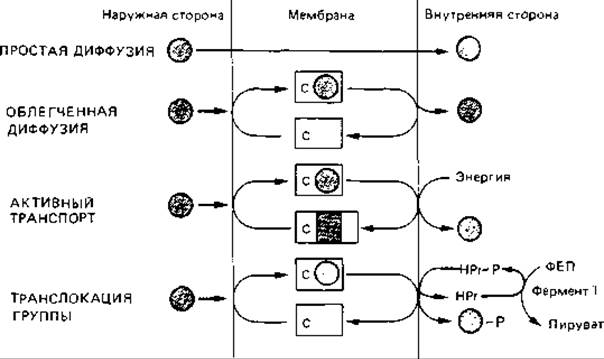
Рис. 7.18. Схема четырех механизмов транспорта веществ в клетку. Розовый кружок - транспортируемый субстрат; с - пермеаза (белок-переносчик); с с серым прямоугольником - энергизованный переносчик; ФЕП - фосфоенолпируват; TБ-термостабильный белок. Пояснения в тексте.
Активный транспорт. Активный транспорт и транслокация группы имеют с облегченной диффузией то общее, что эти процессы происходят с участием субстрат-специфических транспортных белков. Однако в отличие от облегченной диффузии такого рода транспорт требует затраты энергии. При использовании метаболической энергии вещество может накапливаться в клетке против концентрационного градиента. Основное различие между активным транспортом и транслокацией группы заключается в природе молекулы, поступающей внутрь клетки.
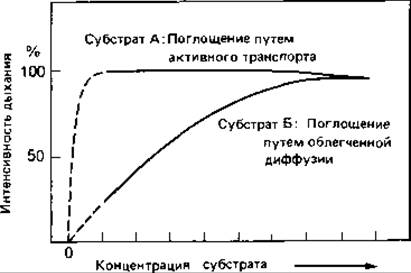
Рис. 7.19. Кривые насыщения при поглощении двух субстратов интактными клетками бактерий [построены по данным о потреблении O2 (интенсивности дыхания)]. Активное и пассивное поглощение субстрата можно различить по форме кривой. Так как субстрат А поглощается путем активного транспорта и накапливается в клетке, дыхание даже при очень малых концентрациях субстрата достигает максимального уровня. Субстрат Б поглощается пассивно, и интенсивность дыхания достигает максимума лишь при относительно высокой концентрации такого субстрата (порядка 10-20 мМ/л).
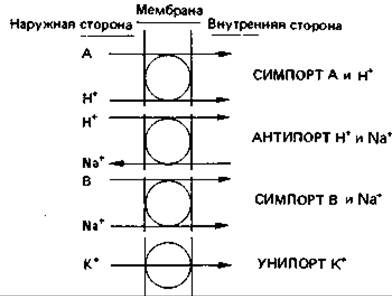
Рис. 7.20. Различные виды активного транспорта, для которых источником энергии служит протонный потенциал ∆р.
При активном транспорте в цитоплазму поступает та же молекула, которая была поглощена из питательной среды. При транслокации группы переносимая молекула в процессе транспорта видоизменяется, например фосфорилируется.
Все теории, объясняющие активный транспорт, включают представление о наличии в мембране специфических транспортных белков. Эти белки получили названия, указывающие на их функцию: пермеазы, транслоказы, белки-транслокаторы, переносчики. Транспортные процессы отличаются друг от друга главным образом тем, что служит для них источником энергии - протонный потенциал Ар (рис. 7.20), АТР или фосфоенолпируват (рис. 7.18).
Для переноса многих веществ, в том числе неорганических и органических ионов, а также сахаров, используется энергия протонного потенциала (см. стр. 243-244). Бактериальные клетки поддерживают протонный потенциал, непрерывно откачивая из клетки протоны и другие ионы (Na+). Для этого в мембране имеются специфические транспортные белки.
Каждый из этих белков имеет совершенно определенную функцию. Есть, например, белок, катализирующий одновременный и однонаправленный перенос одного протона и одной молекулы сахара (лактозы, мелибиозы, глюкозы). В таких случаях говорят о симпорте двух (или нескольких) веществ. Другие транспортные белки катализируют одновременный встречный перенос двух частиц, например одного протона и какого-то другого иона (Na+ или аниона органической кислоты); в этих случаях говорят об антипорте. При переносе сахаров, сопряженном с транспортом ионов, вероятно, всегда используются ионы Н+ или Na+. У прокариот преобладает симпорт с ионами Н+, у эукариот — симпорт с Na+(рис. 7.20).
То, что у бактерий действительно существуют транспортные белки описанного типа, было подтверждено (а) путем очистки и последующего встраивания белка-переносчика в протопласты или в так называемые липосомы и (б) путем выделения мутантов, лишенных соответствующего белка и его специфической функции. Что касается транспорта с использованием энергии протонного потенциала, то это, вероятно, наиболее распространенный механизм активного поглощения субстратов.
Представление об участии специфических белков-переносчиков в транспорте ионов подтверждают данные о действии ряда антибиотиков и синтетических веществ. Речь идет о ионофорах. Это соединения с относительно небольшой молекулярной массой (500-2000), молекулы которых снаружи гидрофобны, а внутри гидрофильны. Обладая гидрофобными свойствами, они диффундируют в липидную мембрану. Из аитибиотиков-ионофоров наиболее известен валиномицин; он диффундирует внутрь мембраны и катализирует транспорт (унипорт) ионов К+, Cs+, Rb+ или NH4+. Поэтому присутствие таких катионов в суспензионной среде приводит к выравниванию заряда по обе стороны мембраны (как бы короткому замыканию) и тем самым к падению протонного потенциала. Другие ионофоры образуют каналы, по которым могут проходить ионы. Существуют также синтетические соединения, повышающие протонную проводимость мембран; наиболее известный переносчик протонов - карбонилцианид-n-трифторметоксифенилгидразон. Он действует как «разобщитель» - нарушает сопряжение синтеза АТР с транспортом электронов, перенося в клетку протоны в обход ATP-синтазы. Изучение мембранного транспорта привело к важным результатам, которые согласуются с хемиосмотической теорией преобразования энергии и подкрепляют ее.
Наряду с транспортными системами, использующими протонный потенциал, существуют также системы, зависимые от АТР. Определенную роль здесь играют периплазматические связующие белки (рис. 2.28). Плазматическая мембрана животных клеток не транспортирует протоны и не создает протонного градиента. Мембранный потенциал, вероятно, поддерживается только ATP-зависимыми насосными механизмами, например натрий-калиевым насосом, а натриевый потенциал в свою очередь доставляет энергию для симпорта питательных веществ вместе с ионами Na+.
Транслокация группы. При транспорте этого типа молекула химически модифицируется; поглощается, например, сахар как таковой, а внутрь клетки он поступает в фосфорилированной форме. Фруктоза, глюкоза, маннитол и родственные вещества поглощаются с помощью фосфотрансферазной системы, зависимой от фосфоенолпирувата. Эта система состоит из неспецифического и специфического компонентов. Неспецифический компонент - это термостабильный белок, который при участии фермента I, находящегося в цитоплазме, фосфорилируется фосфоенолпируватом. Второй компонент - находящийся в мембране индуцибельный фермент II, специфичный для того или иного сахара; он катализирует перенос фосфата с термостабильного белка (ТБ) на сахар во время транспорта последнего через мембрану:
![]()
Фермент II, вероятно, выполняет функцию пермеазы и фосфотрансферазы одновременно (см. рис. 7.18).
В остальном поглощение веществ клетками - процесс очень сложный и пока еще плохо изученный. Многие метаболические эффекты торможения и явления конкуренции между одновременно доступными субстратами связаны, по-видимому, с особенностями регуляторных механизмов, которые проявляются уже в процессах транспорта веществ.
Выход веществ из клетки. О выходе метаболитов в окружающую среду известно существенно меньше, чем о механизмах поглощения веществ клеткой. По-видимому, и выделение их из клетки тоже происходит как при участии транспортных систем, так и путем неконтролируемой диффузии. Вещества выходят из клетки тогда, когда в результате перепроизводства они накапливаются в ней, достигая концентраций, превышающих нормальный уровень. Накопление может быть следствием неполного окисления, нарушения регуляции или процессов брожения.
Транспорт железа. Для транспорта этого макроэлемента микробная клетка обладает специальным механизмом. В анаэробных условиях железо представлено двухвалентным ионом (Fe2+), и его концентрация может достигать 10-1 М/л, так что она не лимитирует рост микроорганизмов. Однако в аэробных условиях при pH 7,0 железо представлено в виде гидроксидного комплекса Fe3+, который почти нерастворим; концентрация ионов трехвалентного железа составляет всего лишь 10-18 М/л. Неудивительно поэтому, что микроорганизмы выделяют вещества, переводящие железо в растворимую форму. Эти вещества - так называемые сидерофоры - связывают ионы Fe3+ в комплекс и в таком виде транспортируют его; речь идет в основном о низкомолекулярных водорастворимых веществах (с мол. массой меньше 1500), связывающих железо координационными связями с высокой специфичностью и высоким сродством (константа устойчивости порядка 1030). По своей химической природе это могут быть феноляты или гидроксаматы. К первым относится энтерохелин; он обладает шестью фенольными гидрокси-группами, и его выделяют некоторые энтеробактерии. Выйдя в окружающую среду, он связывает железо, и образовавшийся ферри-энтерохелин поглощается клеткой. В клетке железо освобождается в результате ферментативного гидролиза ферри-энтерохелина (рис. 7.21).
Многие грибы для той же цели образуют феррихромы; их относят к гидроксаматным сидерофорам. Это циклические гексапептиды, удерживающие трехвалентное железо с помощью трех гидроксаматных групп. Они тоже выделяются из клетки в виде не содержащих железо соединений, связывают в питательной среде железо и в виде феррихромов снова поглощаются. В клетке железо восстанавливается до Fe2+, к которому феррихромы имеют лишь незначительное сродство и поэтому освобождают его. Сходную функцию выполняют ферриоксамины (у актиномицетов), микобактины (у микобактерий) и экзохелины (тоже у микобактерий).
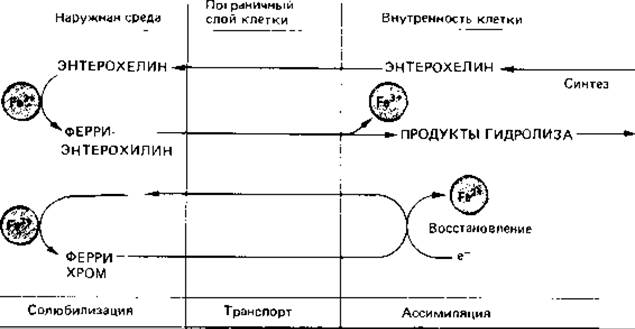
Рис. 7.21. Примеры механизмов переноса железа в клетки микроорганизмов с участием сидерофоров. Вверху - система транспорта с помощью энтерохелина, свойственная многим бактериям; внизу - феррихромная система, имеющаяся у многих грибов.
Микроорганизмы обычно выделяют сидерофоры в питательную среду только тогда, когда железо лимитирует рост. Выделение сидерофоров - следствие дерепрессии их синтеза. В присутствии растворенного, комплексно связанного железа сидерофоры синтезируются лишь в малых количествах и удерживаются в клеточной стенке. В этих условиях они служат только для транспорта железа в клетку.
В связи с этим интересно то, что среди естественных защитных приспособлений высших организмов мы находим «очистку» внутренней среды от железа. Существуют специальные белки, которые так прочно связывают имеющееся железо, что оно становится недоступным для микроорганизмов. Так, например, в белке куриного яйца содержится кональбумин, в молоке, слезной жидкости и слюне - лактотрансферрин, а в сыворотке крови - серотрансферрин. При посеве бактерий на куриный белок они растут только в том случае, если одновременно с инокуляцией вводят ионы железа (в виде цитрата). Таким образом, железо играет важную роль в антагонистических отношениях между высшими организмами и бактериями. Борьбу выигрывает тот партнер, который вырабатывает вещество, прочнее связывающее железо.