Общая микробиология - Шлегель Г. 1987
Использование неорганических доноров водорода: аэробные хемолитотрофные бактерии
Окисление аммиака и нитрита (нитрификация)
Многие группы почвенных и водных бактерий могут использовать в качестве доноров водорода или электронов неорганические соединения или ионы (ионы аммония, нитрита, сульфида, тиосульфата, сульфита и двухвалентного железа), а также элементарную серу, молекулярный водород и СО, т. е. способны получать в результате их окисления восстановительные эквиваленты и энергию для синтетических процессов. Получение энергии происходит, как правило, в результате дыхания с О2 как конечным акцептором водорода. Лишь немногие из относящихся к этой группе бактерий способны расти за счет «анаэробного дыхания», используя в качестве акцепторов водорода нитрат, нитрит, закись азота и т. п. Такой образ жизни с использованием неорганического донора водорода называют хемолитотрофным.
Большинство бактерий с таким типом метаболизма используют СО2 в качестве единственного или главного источника клеточного углерода. Они являются поэтому автотрофами (хемолитоавтотрофами). Почти все аэробные хемолитоавтотрофные бактерии, до сих пор изученные в этом отношении, ассимилируют углерод СО2 через рибулозобисфосфатный цикл1. Механизм такого типа фиксации СО2 будет рассмотрен в конце этой главы (разд. 11.5).
Для некоторых хемолитотрофных бактерий такой образ жизни является облигатным, другие же - факультативные хемолитотрофы, т. е. способны также и к хемоорганогетеротрофному росту. Многие хемолитоавтотрофные бактерии благодаря своей высокой специализации занимают монопольное положение. Окисление аммиака, нитрита и неорганических соединений серы в природе в первую очередь связано с деятельностью нитрифицирующих и серуокисляющих бактерий.
1 Исключение составляет аэробная водородокисляющая бактерия Hydrogenobacter thermophilus - Прим. ред.
При аэробном или анаэробном разложении азотсодержащих органических веществ происходит выделение азота в форме аммиака. Давний опыт учит, что при компостировании навоза образуется селитра. Селитряные выцветы, появляющиеся на каменной облицовке навозных ям, в средние века использовали для приготовления пороха. Согласно старым руководствам, на селитряных заводах из смеси земли, известняка и азотсодержащих органических веществ закладывали специальные гряды. Эти гряды поливали мочой и кровью, следя за тем, чтобы они хорошо аэрировались. Аммиак, освобождавшийся при микробном разложении органического материала, диффундировал в верхний земляной покров и окислялся там под воздействием кислорода воздуха до нитрата. Этот покрывающий верхний слой земли служил исходным материалом для получения селитры; его вымачивали в воде, а полученный раствор затем выпаривали.
Превращение аммиака (аммония) в нитрат - нитрификация - как в почве, так и в воде осуществляется нитрифицирующими бактериями. Однако нет такой бактерии, которая бы прямо превращала аммиак в нитрат. В его окислении всегда участвуют две группы бактерий: одни окисляют аммиак, образуя нитрит, а другие окисляют нитрит в нитрат.
Наиболее известные виды нитрифицирующих бактерий - это Nitrosomonas еuгораеа и Nitrobacter winogradskyi (табл. 11.1). Проведенные недавно исследования показали, что важнейшими нитрификаторами в почвах сельскохозяйственных угодий являются виды рода Nitrosolobus, а не Nitrosomonas, как считалось ранее. Обе группы родов строго специализированы в отношении приведенных в табл. 11.1 реакций. Бактерии, окисляющие аммиак, поставляют субстрат для бактерий, окисляющих нитрит. Поскольку высокие концентрации аммиака в щелочных почвах оказывают на Nitrobacter токсическое действие, Nitrosomonas, используя аммиак и образуя кислоту (т. е. переводя катион в анион), тем самым улучшает и условия существования для Nitrobacter.
Таблица 11.1. Нитрифицирующие бактерии
|
Бактерии, окисляющие аммиак (Nitroso-) |
Бактерии, окисляющие нитрит (Nitro-) |
|
NH+4 + 11/2O2 - NO-2 + 2H+ + H2O |
NO-2 + 1/2O2→ NO3 |
|
Nitrosomonas еигораеа |
Nitrobacter winogradskyi |
|
Nitrosococcus oceanus |
Nitrobacter agilis |
|
Nitrosospira briensis |
Nitrospina gracilis |
|
Nitrosolobus multiformis |
Nitrococcus mobilis |
Нитрификаторы - это грам-отрицательные бактерии, принадлежащие к семейству Nitrobacteraceae. У Nitrosomonas еurораеа клетки овальные с полярно расположенными жгутиками. В морях за окисление аммиака ответствен, по-видимому, Nitrosococcus oceanus (рис. 2.22). Нитрифицирующие бактерии можно выращивать на чисто минеральных средах, однако в таких условиях они растут медленно (время генерации 10-20 ч). До последнего времени нитрификаторов считали облигатными хемолитоавтотрофами, поскольку они не используют добавляемые к питательным средам органические субстраты. Однако сейчас на этот счет высказывается все больше сомнений, и вопрос детально изучается.
Оказалось, например, что Nitrobacter winogradskyi способен использовать добавленный к минеральной среде ацетат для синтеза некоторых веществ клетки (белков, поли-ß-гидроксимасляной кислоты).
Этапы окисления аммиака. При окислении, по-видимому, образуется ряд промежуточных продуктов:
![]()
Первый этап окисления является эндергонической "реакцией, которую катализирует монооксигеназа. Атом кислорода в NH2OH доставляется молекулярным кислородом. Второй этап идет при участии гидроксиламин-оксидоредуктазы. При окислении нитрита электроны переносятся на цитохром a1. Только стадии окисления гидроксиламина в нитрит и нитрита в нитрат являются энергетически полезными.
Роль процессов нитрификации в почве. В хорошо аэрируемой почве ионы NH+4, освобождающиеся при минерализации азотсодержащих веществ, подвергаются быстрому окислению. Перевод катиона в анион ведет к подкислению почвы и тем самым к повышению растворимости минералов (солей калия, магния, кальция и фосфорной кислоты). Поэтому в нитрифицирующих микроорганизмах видели ранее важный фактор плодородия почв. Однако теперь эти представления изменились. Выяснилось, что ионы аммония задерживаются в почве гораздо лучше нитрата, особенно если они адсорбируются на глинистых минералах и более или менее прочно связываются с частицами гумуса; нитрат же легко вымывается. В связи с этим появилась тенденция к ограничению нитрификации в почвах сельскохозяйственных угодий. Ведутся поиски веществ, способных специфически подавлять рост нитрифицирующих бактерий и служить своего рода «стабилизаторами» почвенного азота [к таким веществам относится, например, 2-хлор-6-(трихлорметил)-пиридин].
Следует учесть, что рост и метаболизм автотрофных нитрифицирующих бактерий протекает оптимально лишь в области pH от 7 до 8. Диапазон pH, в котором происходит полная нитрификация от аммиака до нитрата, очень узок, поскольку и свободный аммиак (при высоких значениях pH), и азотная кислота (при низких значениях pH) оказывают токсическое действие на Nitrobacter. Известно, что концентрации свободного NH3 и свободной HNO2зависят от pH среды.
Нитрифицирующие бактерии косвенно участвуют в разрушении разного рода сооружений, для которых строительным материалом служат известь и цемент (т. е. различных зданий, автострад и т. п.). Это связано с тем, что нитрифицирующие бактерии окисляют аммиак, присутствующий в атмосфере или выделяющийся из фекалий животных, до азотной кислоты.
Гетеротрофная нитрификация. Пока не установлено точно, действительно ли автотрофные нитрификаторы занимают монопольное положение в природе или же превращение аммиака в нитрат происходит также при участии гетеротрофных бактерий и грибов. В чистой культуре лишь некоторые штаммы Arthrobacter способны к образованию нитрита из азотсодержащих веществ. Некоторые грибы обладают способностью окислять аминный азот или аммиак до нитрата. Однако гетеротрофный процесс в отличие от автотрофной нитрификации не связан с ростом клеток и продукцией биомассы. Речь при этом идет, вероятно, о какого-то рода соокислении аммиака и органических субстратов. Кроме того, скорость нитрификации у гетеротрофных бактерий в 103-104 раз меньше, чем у автотрофных. Поэтому нельзя считать, что гетеротрофные нитрификаторы серьезно «подрывают» монопольное положение автотрофных нитрифицирующих организмов. Однако даже незначительная нитрификация, осуществляемая гетеротрофами, позволяет объяснить, например, тот факт, что и на кислых почвах (чайные плантации, хвойные леса), где у автотрофных нитрифицирующих микроорганизмов нет шансов выжить, все же происходит образование нитрата.
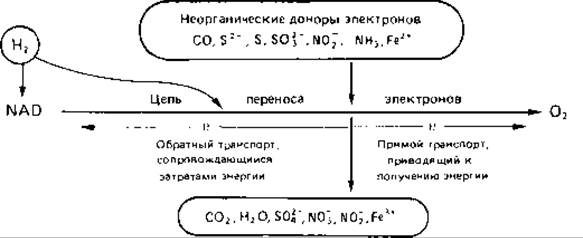
Обратный транспорт электронов и урожай клеток. Автотрофные бактерии, окисляющие аммиак, нитрит, соединения серы или железо, находятся в энергетическом отношении в крайне невыгодных условиях. Их субстраты обладают сильно положительным окислительно-восстановительным потенциалом. Нормальные потенциалы Е0' составляют для NH+4/NH2OH + 899 мВ, для NO-3 /NO-2 + 420 мВ, для NO-2/NH2OH + 66 мВ, для Fe3 + /Fe2++ 770 мВ, в то время как для пары NAD/NADH2 окислительно-восстановительный потенциал равен — 320 мВ. Окисление названных выше неорганических субстратов не может быть прямо связано с восстановлением NAD. Однако NADH2 необходим для восстановления СO2 в рибулозобисфосфатном цикле. Судя по некоторым экспериментальным данным, электроны, освобождающиеся при окислении упомянутых неорганических субстратов, поступают в дыхательную цепь на уровне цитохрома с или цитохрома а. Поскольку фосфорилирование при этом может происходить лишь на одном-единственном этапе окисления, выигрыш в энергии соответственно невелик. Часть этой энергии затрачивается на то, чтобы «оттеснить» электроны, поступающие на участок цитохромов, по дыхательной цепи назад, на уровень пиридиннуклеотидов, и восстановить последние. Таким образом, обратный перенос электронов является для перечисленных выше бактерий обязательным механизмом, необходимым для получения восстановительных эквивалентов, используемых затем в процессах синтеза (см. стр. 246).
Таблица 11.2. Сравнение количеств первичного источника энергии, необходимых различным автотрофным и органотрофным микроорганизмам для образования 1 г сухой клеточной массы
|
Thiobacillus ferrooxidans |
156 г Fe2+ |
Alcaligenes eutrophus |
0,5 г H2 |
|
Thiobacillus neapolitanus |
30 г S2O2-3 |
Escherichia coli |
2 г глюкозы |
|
Nitrosomonas |
30 г NH3 |
Дрожжи |
1 г нефти |
Низкий выход клеток у бактерий, окисляющих упомянутые выше неорганические субстраты, обусловлен тем, что они получают таким способом мало энергии. Для синтеза одного грамма сухой клеточной массы им приходится использовать гораздо большие количества различных веществ, чем другим организмам (табл. 11.2). Кроме того, большинство названных бактерий катализирует так называемое «холостое окисление», т. е. окисление субстратов без одновременного синтеза клеточного вещества. Не удивительно поэтому, что различные превращения в почве и воде осуществляются сравнительно малым количеством бактерий.