Общая микробиология - Шлегель Г. 1987
Постоянство, изменение и передача признаков
Синтез белка и генетический код
Всякое живое существо по большинству своих признаков сходно со своими предками. Сохранение специфических свойств, т. е. постоянство признаков в ряду поколений, называют наследственностью. Изучением передачи признаков и закономерностей их наследования занимается генетика. Каждому признаку в качестве носителя информации соответствует определенный ген. Еще во времена классической генетики исследователи пришли к выводу, что гены находятся в клеточном ядре. Тогда же было установлено, что они должны располагаться в линейном порядке. Долгое время считали, что наследственная информация связана с белковыми компонентами нуклеоплазмы. Лишь после успешных экспериментов по передаче наследственных признаков с помощью ДНК (см. разд. 15.3.4) генетики пришли к убеждению, что именно ДНК, входящая в состав хромосом у всех организмов, служит материальным носителем наследственной информации. Сначала на насекомых, а затем на микроорганизмах было показано, что проявление признаков зависит от активности ферментов. У микроорганизмов ферменты можно было связать с конкретными признаками, поддающимися точному биохимическому определению. Гипотеза один ген - один фермент гласит, что определенный ген содержит информацию, необходимую для синтеза определенного фермента (позднее была принята более точная формулировка: каждый структурный ген кодирует определенную полипептидную цепь). Изменение гена вследствие мутации приводит либо к утрате фермента, либо к изменению его свойств, а тем самым и к изменению признака. Гены выявляются только благодаря мутациям. Генетический анализ основан прежде всего на изучении различий в признаках, определяемых альтернативными формами (аллелями) того или иного гена. Поэтому исследование различных генетических проблем ведется на мутантах.
Микроорганизмы - чрезвычайно удобные объекты для генетического анализа, ибо опыты можно проводить в короткие сроки на огромном числе особей и они не требуют много места. Однако эти преимущества были оценены не сразу: бактерии стали использоваться в генетических исследованиях лишь тогда, когда был преодолен ряд давних предубеждений.
Репликация ДНК. У бактерий, так же как и у высших организмов, носителем генетической информации служит ДНК. Рассматривая структуру клетки, мы уже говорили о том, что бактериальная ДНК представляет собой двойную спираль, замкнутую в кольцо. Сразу же возникает вопрос: как сохраняется наследственная информация при росте и размножении клеток? Перед их делением происходит идентичная редупликация, или репликация, генов. Этот процесс можно удовлетворительно объяснить, исходя из модели структуры ДНК, предложенной Уотсоном и Криком, и из механизма удвоения ДНК, теперь уже известного (см. с. 36). Две цепи двойной спирали ДНК комплементарны друг другу. На каждой цепи из структурных элементов ДНК-дезоксирибонуклеозидтрифосфатов - синтезируется новая цепь; при этом с каждым из оснований спаривается комплементарное ему основание, так что каждая из двух новых цепей опять-таки будет комплементарна родительской цепи. Обе новые двойные спирали состоят из одной родительской и одной вновь синтезированной цепи. Эта точная репликация ДНК гарантирует сохранение генетической информации.
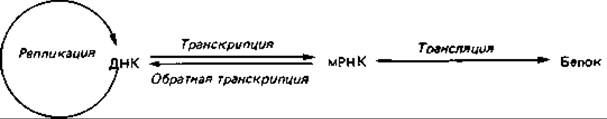
Транскрипция ДНК. Возникает еще один вопрос: каким образом содержащаяся в генах информация определяет специфическую активность и другие свойства ферментов и как она преобразуется в аминокислотную последовательность ферментного белка? ДНК, будучи носителем наследственной информации, тем не менее сама не служит матрицей для синтеза полипептидов. Биосинтез белков происходит на рибосомах, которые непосредственно с ДНК не соприкасаются. Передачу записанной в ДНК информации к местам синтеза белка осуществляет матричная, или информационная, рибонуклеиновая кислота (мРНК). Она состоит из одной цепи и очень напоминает одиночную цепь ДНК с тем отличием, что тимин (Т) ДНК в РНК заменен урацилом (U). мРНК синтезируется на одной из цепей ДНК, причем механизм этого процесса сходен с механизмом репликации ДНК. Образование мРНК начинается на 5'-ОН-конце, и по последовательности оснований ее цепь комплементарна цепи ДНК. Таким образом, при синтезе мРНК просто копируется нуклеотидная последовательность ДНК. Этот процесс называют транскрипцией и противопоставляют его трансляции - переводу нуклеотидной последовательности в последовательность аминокислот:
Генетический код. Каждый ген представлен определенным участком молекулы ДНК. Специфическая информация, содержащаяся в гене, определяется последовательностью оснований в цепи ДНК. «Алфавит», с помощью которого записана эта информация ДНК. включает четыре «буквы» - основания аденин (А), гуанин (G), тимин (Т) и цитозин (С). В мРНК тимин заменен урацилом (U).
Специфичность ферментных белков, синтез которых контролируют гены, определяется последовательностью аминокислот в полипептидных цепях. Эта же последовательность определяет и пространственную структуру белка, так называемую конформацию (вторичную, третичную и четвертичную структуру).
Для перевода с языка нуклеиновых кислот на язык аминокислот служит специфический код. Каждая аминокислота определяется группой из трех соседних нуклеотидов - триплетом, или кодоном. Та или иная последовательность триплетов в нуклеиновой кислоте однозначно определяет последовательность аминокислот в полипептидной цепи; последняя, таким образом, представляет собой колинеарное отображение нуклеиновой кислоты. В триплетах возможны 64 различные комбинации нуклеотидов (табл. 15.1). Если бы каждая из 20 аминокислот кодировалась лишь одним-единственным триплетом, то 44 возможные комбинации остались бы неиспользованными. Но оказалось, что многие аминокислоты кодируются двумя или большим числом различных триплетов. Некоторые триплеты имеют особый смысл-они означают «начало» или «конец» полипептидной цепи. Триплеты считываются как 1, 2, 3; 1, 2, 3 и т.д. от начала молекулы мРНК.
Таблица 15.1. Генетический код. Триплеты, или кодоны, UAA (ochre) UAG (amber) и UGA определяют конец синтеза и отделение растущей полипептидной цепи от рибосомы
|
Триплет |
Аминокислота |
Триплет |
Аминокислота |
Триплет |
Аминокислота |
Триплет |
Аминокислота |
|
UUU |
Phe |
UCU |
Ser |
UAU |
Tyr |
UGU |
Cys |
|
UUC |
Phe |
UCC |
Ser |
UAC |
Tyr |
UGC |
Cys |
|
UUA |
Leu |
UCA |
Ser |
UAA |
«ochre» |
UGA |
— |
|
UUG |
Leu |
UCG |
Ser |
UAG |
«amber» |
UGG |
Trp |
|
CUU |
Leu |
CCU |
Pro |
CAU |
His |
CGU |
Arg |
|
CUC |
Leu |
CCC |
Pro |
CAC |
His |
CGC |
Arg |
|
CUA |
Leu |
ССА |
Pro |
CAA |
Gln |
CGA |
Arg |
|
CUG |
Leu |
CCG |
Pro |
CAG |
Gln |
CGG |
Arg |
|
AUU |
Ileu |
ACU |
Thr |
AAU |
Asn |
AGU |
Ser |
|
AUC |
eu |
АСС |
Thr |
AAC |
Asn |
AGC |
Ser |
|
AUA |
Ileu |
АСА |
Thr |
AAA |
Lys |
AGA |
Arg |
|
AUG |
Met |
ACG |
Thr |
AAG |
Lys |
AGG |
Arg |
|
GUU |
Val |
GCU |
Ala |
GAU |
Asp |
GGU |
Gly |
|
GUC |
Val |
GCC |
Ala |
GAC |
Asp |
GGC |
Gly |
|
GUA |
Val |
GCA |
Ala |
GAA |
Glu |
GGA |
Gly |
|
GUG |
Val |
GCG |
Ala |
GAG |
Glu |
GGG |
Gly |
|
Ala - |
аланин |
Gly - глицин |
Pro |
- пролин |
|||
|
Arg - |
аргинин |
His - гистицид |
Ser |
- серин |
|||
|
Asn - |
аспарагин |
Ile - изолейцин |
Thr |
- треонин |
|||
|
Asp - |
аспарагиновая кислота |
Leu - лейциин |
Trp |
- триптофан |
|||
|
Cys - |
цистеин |
Lys - лизин |
Tyr |
- тирозин |
|||
|
GLn - |
глутамин |
Met - метионин |
Val |
- валин |
|||
|
Glu - |
глутаминовая кислота |
Phe - фенилаланин |
|||||
Трансляция мРНК: синтез белка. Аминокислоты соединяются в полипептидную цепь в порядке, определяемом триплетами мРНК. В этом процессе участвуют мРНК, транспортные РНК (тРНК). рибосомы, ряд ферментов, АТР и другие факторы. Сначала аминокислоты при участии АТР активируются с образованием аминоацил-АМР:
Аминокислота + АТР → Аминоацил-АМР + РРі
От АМР аминоацильная группа переносится на концевой нуклеотид тРНК. Активация и присоединение аминокислоты к соответствующей тРНК осуществляются с помощью специфического фермента-аминоацил-тРНК-синтетазы, которая распознает, с одной стороны, аминокислоту, а с другой - соответствующую тРНК. Имеется 20 различных аминоацил-тРНК-синтетаз, по одной для каждой аминокислоты. Как уже упоминалось, некоторым аминокислотам соответствует не один, а несколько кодонов (т. е. генетический код, как говорят, «вырожденный»); для каждой из таких аминокислот имеется несколько тРНК. Различные тРНК, предназначенные для одной и той же аминокислоты, называют также изоакцепторными тРНК. Таким образом, соответствующая синтетаза может присоединять аминокислоту к нескольким изоакцепторным тРНК. В молекуле тРНК имеется участок, комплементарный кодону мРНК (антикодон). Соединение аминокислот происходит на рибосомах (рис. 15.1). Рибосома перемещается вдоль мРНК, начиная с 5'-ОН-конца, и при каждом ее перемещении на один триплет очередная аминокислота устанавливается транспортной тРНК в нужное положение и присоединяется своей аминогруппой к карбоксильной группе предшествующей аминокислоты (образуется пептидная связь). Так растет полипептидная цепь по мере продвижения рибосомы вдоль мРНК. По-видимому, одновременно происходит закручивание этой цепи и свертывание ее в клубок, определяемое последовательностью аминокислот и природой их боковых цепей (гидрофобные и гидрофильные группы), и в результате возникает структура, обусловливающая специфические свойства и функцию данного белка. К мРНК обычно прикрепляется несколько рибосом, так что на одной и той же матрице одновременно синтезируется несколько полипептидных цепей. Такой комплекс одной мРНК с рибосомами называют полисомой. На конце мРНК находится кодон, от которого зависит отделение сформированной полипептидной цепи от рибосомы (UAA, UAG или UGA).
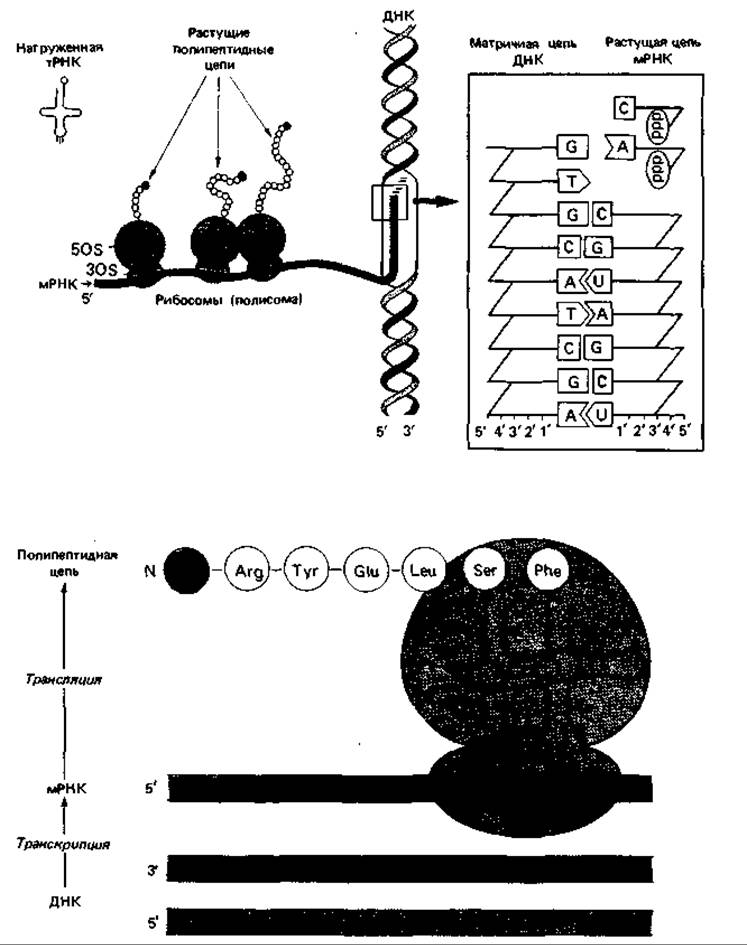
Рис. 15.1. Биосинтез белка. Перенос генетической информации осуществляется в два этапа. Сначала на матричной цепи ДНК образуется мРНК. Затем во время перемещения рибосомы вдоль мРНК (на схемах - слева направо) различные тРНК подводят к ней аминокислоты и устанавливают их в положение, определяемое триплетами мРНК. Аминокислоты соединяются между собой пептидными связями.
Таким образом, нуклеотидная последовательность ДНК представляет собой закодированную «инструкцию», определяющую (при посредстве мРНК) структуру специфического белка. Представление о передаче информации от ДНК через РНК на белок называют «центральной догмой» молекулярной биологии. Таким путем происходит перенос информации у всех организмов, у которых генетическим материалом служит ДНК. Этот универсальный процесс передачи информации при репликации ДНК, транскрипции и трансляции представлен на приведенной выше схеме (стр. 435) красными стрелками. Эта схема применима к эукариотам, прокариотам и ДНК-вирусам.
Среди РНК-вирусов есть такие, у которых РНК реплицируется прямо на матрице РНК. Однако у некоторых онкогенных (опухолеродных) РНК-вирусов вначале происходит синтез ДНК, контролируемый РНК, т. е. РНК служит матрицей для синтеза ДНК. Таким образом, информация, содержащаяся в вирусной РНК, передается на ДНК путем обратной транскрипции (при помощи фермента обратной транскриптазы; см. упомянутую выше схему). Этот фермент можно выделить из клеток опухолей, вызываемых РНК-вирусами. Он находит применение в генной инженерии (см. разд. 15.3.6). Если, например, в качестве носителя информации выделяют не фрагмент ДНК, а соответствующую мРНК, то последняя должна быть «переписана» в ДНК, которая и встраивается в плазмиду. При помощи обратной транскриптазы удается получить нужную ДНК in vitro.
У прокариотических организмов обратная транскрипция не найдена.