Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Белки
Белки в роли ферментов: лизоцим
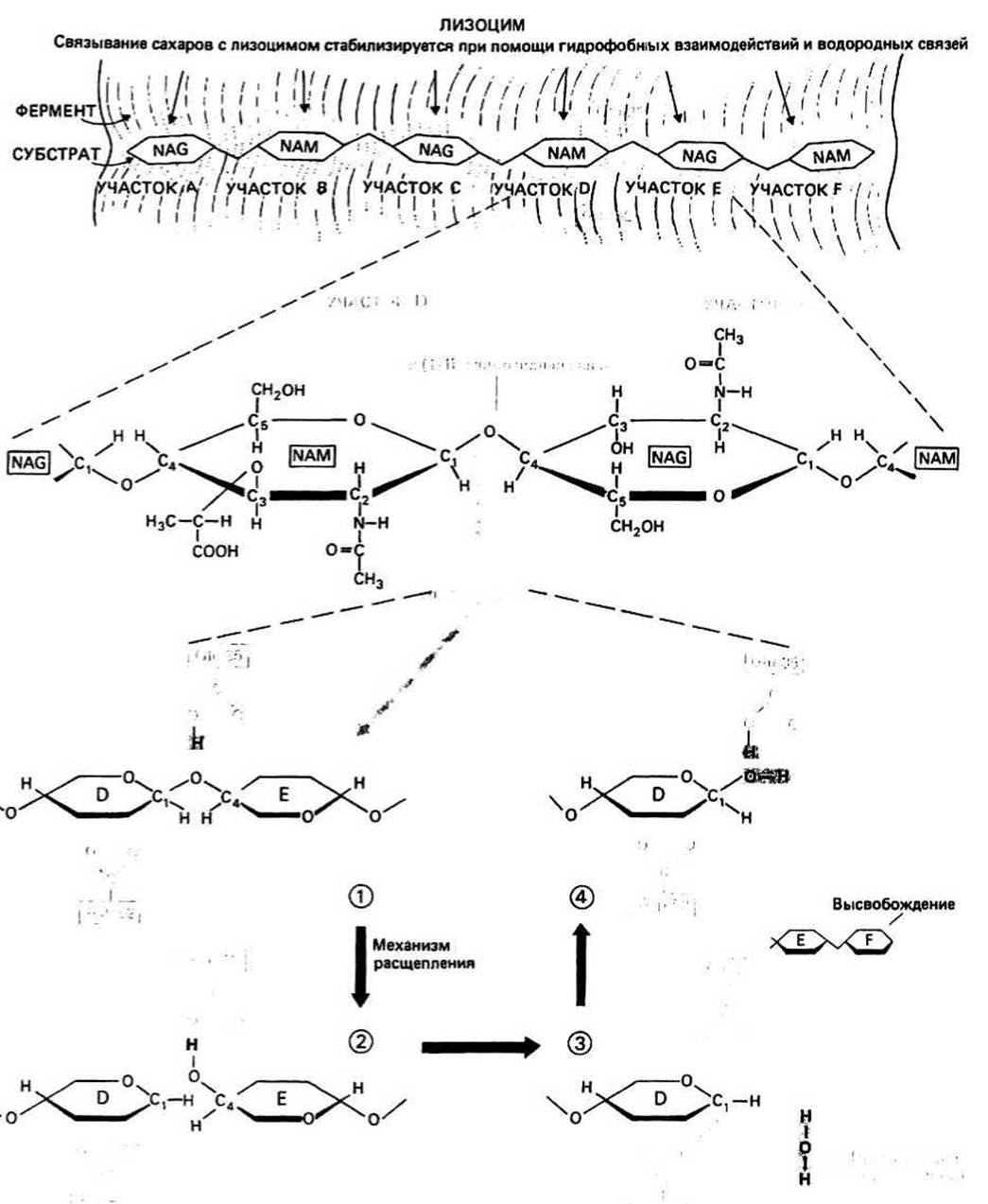
Рис. 14.1.
Лизоцим — это фермент, способный разрушать определенные бактериальные клетки, расщепляя полисахаридные цепи клеточной стенки. Лишенная жесткой клеточной стенки, бактерия разрывается под действием осмотического шока, вызываемого быстрым проникновением воды внутрь клетки.
Полисахарид клеточной стенки представляет собой полимер, в котором чередуются остатки сахаров двух типов — N-ацетилглюкозамина (NAG) и N-ацетилмурамовой кислоты (NAM). Сахара, имеющие ß-конфигурацию относительно аномерного С1 -атома, образуют полимерную цепь с помощью гликозидных связей между С1 -атомом одного сахарного кольца и С4-атомом следующего (гл. 31).
Структура лизоцима. Лизоцим из белка куриного яйца состоит из одной полипептидной цепи, насчитывающей 129 остатков и имеющей четыре дисульфидных мостика (гл. 10). Фермент может связываться
Расщепление полисахарида осуществляется путем гидролиза гликозидной связи между сахарными кольцами, располагающимися в участках D и Е. Анализ кристаллографических данных позволил предположить, что непосредственно перед и во время гидролиза сахарное кольцо в участке D имеет не обычную конформацию кресла, а конформацию полукресла, характеризующуюся тем, что в ней пять из шести атомов, образующих сахарное кольцо, лежат практически в одной плоскости (рис. 14.3)
Центральную роль в функционировании лизоцима играют остатки глутаминовой кислоты 35 и аспарагиновой кислоты 52. Боковые цепи этих остатков располагаются близко к гликозидной связи (примерно на расстоянии 0,3 нм) между сахарными кольцами, локализованными в участках D и Е. Glu 35 находится в неполярном окружении, и поэтому его карбоксильная группа остается протонированной (т. е. с ингибиторами, химически сходными с полисахаридом клеточной стенки. С помощью рентгеноструктурного анализа Филлипсом и др. была определена пространственная структура белка как такового и белка, закристаллизованного вместе с ингибитором. Оказалось, что лизоцим состоит из двух доменов, образующих щель, в которой находится активный центр, способный связать гексосахарид, причем для связывания каждого из шести сахарных колец на ферменте имеется свой участок (эти участки обозначаются буквами А, В, С, D, Е и F) (рис. 14.2). находится в форме —СООН). Окружение Asp52, наоборот, полярно, поэтому карбоксильная группа этого остатка депротонирована (т. е. находится в форме —СООН).
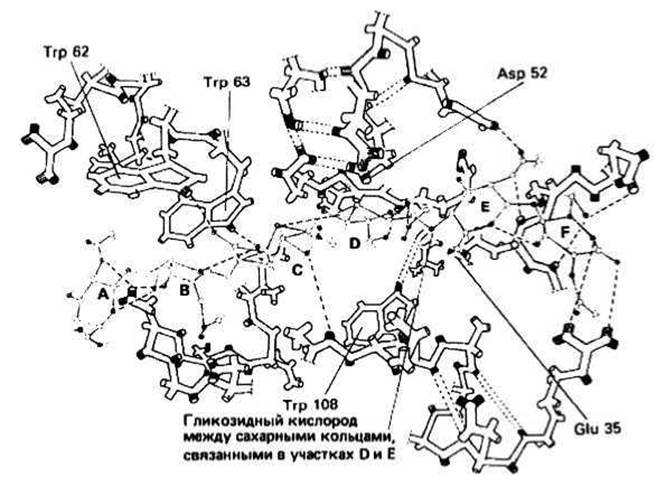
Рис. 14.2. Структура молекулы лизоцима в области активного центра. Коричневым цветом изображен связанный в активном центре субстрат (NAG-NAM)3. NH- и СО-группы соответственно выделены серым и черным цветами. Водородные связи изображены пунктирными прямыми. Обратите внимание на близость колец двух триптофановых остатков Тrf 62 и Тrр 63 к сахарам, располагающимся в участках А и В. Благодаря вандерваальсовым контактам и образованию водородных связей с этими остатками осуществляется дополнительная фиксация субстрата в активном центре.
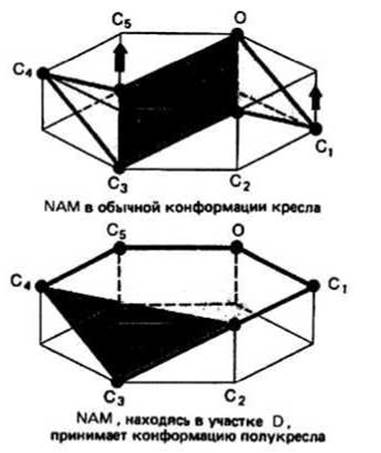
Рис. 14.3.
Реакцию гидролиза можно подразделить на несколько этапов.
1. —СООН-группа остатка Glu 35 предоставляет свой протон гликозидному кислороду, что приводит к разрыву связи между этим атомом кислорода и С1-атомом сахарного кольца, располагающегося в участке D. Получившийся в результате фрагмент исходного полисахарида, включающей в себя сахарные кольца, которые находятся в участках Е и F, является продуктом и может освободиться из комплекса с ферментом.
2. Сахарное кольцо, располагающееся в D-участке, имеет искаженную конформацию, соответствующую конформации переходного состояния. При этом Сrатом оказывается положительно заряженным. Углеродный атом в таком состоянии называется карбоний-ионом. Он стабилизируется с помощью отрицательного заряда близко расположенного остатка Asp 52.
3. Гидроксильный ион (ОН), донором которого служит молекула воды из окружающей среды, присоединяется к карбоний-иону, после чего второй фрагмент расщепленного полисахарида становится продуктом реакции. Одновременно из-за связывания иона водорода (Н+) протонируется карбоксильная группа Glu 35, переходя в форму —СООН.
4. Теперь фермент находится в первоначальном состоянии и готов осуществлять следующую реакцию расщепления полисахарида.
Этот пример позволяет проследить некоторые общие принципы ферментативного катализа
1. Увеличение энергии субстрата за счет искажения структуры сахарного кольца NAM, находящегося в участке D.
2. Наличие необычного окружения Glu 35, обусловливающее появление реакционноспособного протона.
3. Правильная ориентация протона в Glu 35, необходимая для атаки гликозидной связи.
4. Уменьшение свободной энергии переходного со стояния за счет стабилизации карбоний-иона карбок сильной группой остатка Asp 52.