Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Внутриклеточный транспорт белков
Перенос белков через ядерную мембрану
Ядро - наиболее крупная (диаметром около 10 мкм) видимая в оптический микроскоп органелла эукариотической клетки. Оно отделено от остальной клетки оболочкой, состоящей из внутренней и внешней ядерных мембран (рисунок 13). Область между двумя ядерными мембранами называется перинуклеарным пространством. Внешняя ядерная мембрана усыпана рибосомами и переходит в шероховатый эндоплазматический ретикулум.
Внутренняя ядерная мембрана выстлана специальными белками, которые служат для закрепления ядерных структур (ядерная ламина). В ядре расположена почти вся ДНК клетки, и здесь происходит её репликация и экспрессия.
Эти процессы обслуживаются множеством белковых факторов, которые, как и все белки, синтезируются в цитозоле клетки, а затем доставляются в ядро. А матричная РНК - продукт транскрипции ДНК - после процессинга выходит из ядра в цитозоль, равно как и рибосомальные субъединицы после синтеза в ядрышке доставляются из ядра в цитозоль клетки.
Этот интенсивный обмен веществ между цитозолем и нуклеоплазмой осуществляется и управляется ядерными порами - сложными мультибелковыми комплексами, в которые входит более чем 50 белков, которые называют нуклеопоринами, и которые формируют круг из восьми идентичных субъединиц (рисунок 78).
На схеме бокового разреза (рисунок 78(6)) видно, что белки в ядерной поре располагаются в виде стопки из трёх колец белков и крепятся в ядерной мембране с помощью трансмембранных белков. От внутреннего кольца к центру канала поры отходят радиальные "спицы». В центре поры располагается центральный белок-транспортёр, который называется клапан (plug), или пробка, и который выполняет функцию ворот (gate), закрывающих проход через пору.
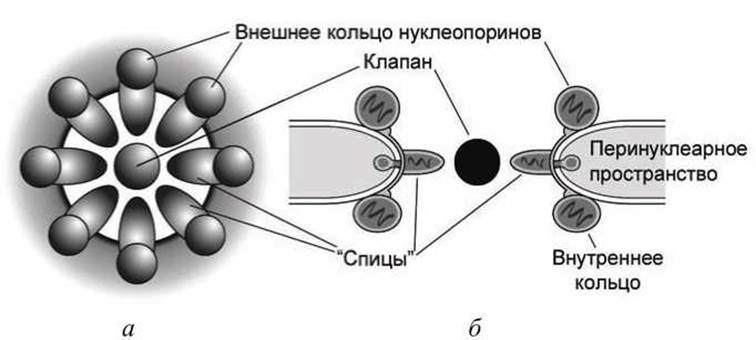
Рисунок 78 - Схема строения ядерной поры: а - фронтальный вид; б - боковой разрез
Определяющую роль в осуществлении управляемого транспорта через ядерную пору играет ГТФаза Ran. Белок Ran, как и все ГТФазные переключающие белки, существует в двух формах (рисунок 79):
(1) активная ("вкл") форма с присоединённой молекулой ГТФ;
(2) неактивная ("выкл") форма со связанной молекулой ГДФ.
ГТФазная активность таких белков-переключателей приводит к медленному гидролизу ГТФ в ГДФ, "выключая" белок. Последующая замена ГДФ на ГТФ, восстанавливающая активную форму, происходит ещё более медленно.
Процессы переключения ГТФазы между двумя конформациями управляются вспомогательными белками: белком Ran-GAP (GTPase-Accelerating Protein), ускоряющим гидролиз ГТФ, и белком Ran-GEF (Guanine nucleotide Exchange factor), стимулирующим замену ГДФ на ГТФ.
В случае транспорта через ядерную пору белки Ran-GEF сконцентрированы в ядре, а белки Ran-GAP - в цитозоле, поэтому в нуклеоплазме ГТФаза Ran находится преимущественно в активном (Rаn:ГТФ)-состоянии, а в цитозоле - в неактивном Rап:ГДФ)-состоянии.
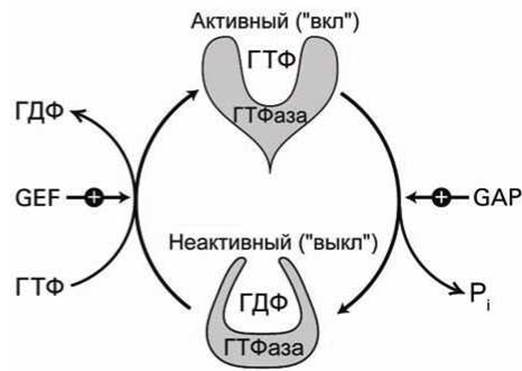
Рисунок 79 - Циклирование белка ГТФаза Ran между активным и неактивным состояниями
Схема импорта белков в ядро, регулируемая белками Ran представлена на рисунке 80. Белок-транспортёр распознает и присоединяет к себе нацеливающую последовательность (ядерную импортную последовательность) белка-груза.
Такой комплекс двух белков диффундирует через ядерную пору в нуклеоплазму, где много активных ГТФаз Ran. Активная ГТФаза замещает белок-груз, и сама связывается с белком-транспортёром. Груз оказывается в ядре.
Комплекс белка-транспортёра с активной ГТФазой диффундирует через ядерную пору наружу в цитоплазму, где находятся белки Ran-GAP. Белок Ran-GAP "выключает" ГТФазу, и она отсоединяется от транспортного белка, который теперь готов снова "загружаться" и транспортировать белки в ядро.
Неактивная ГТФаза диффундирует в ядро, где её активизирует (заменяет ГДФ на ГТФ) белок Ran-GEF.
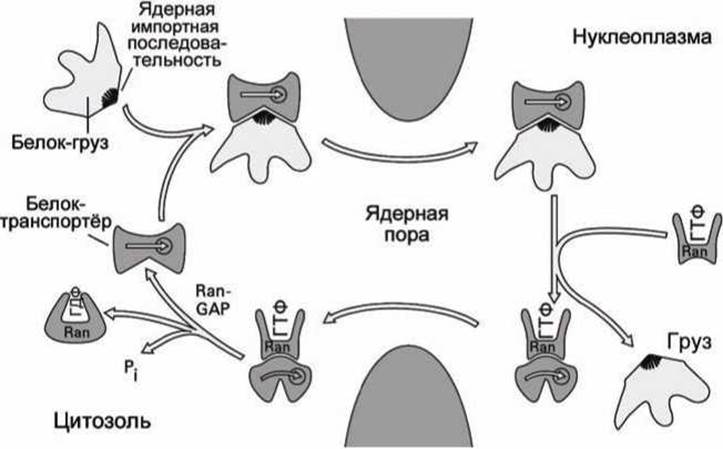
Рисунок 80 - Импорт белков в ядро
Аналогично проходит процесс экспорта белков (рисунок 81), с тем только отличием, что белок-транспортёр сможет распознать нацеливающую последовательность в белке-грузе только, если сам белок-транспортёр предварительно образует двойной комплекс с ГТФазой. Через пору наружу может выйти только тройной комплекс (ГТФаза:Транспортёр:Груз). В цитозоле вспомогательный белок Ran-GAP стимулирует гидролиз ГТФ, ГТФаза "выключается", тройной комплекс распадается, и белок-груз оказывается в цитозоле.
Затем белок-транспортёр и неактивный белок Ran диффундируют в нуклеоплазму, белок Ran-GEF активизирует ГТФазу, и процесс может начаться снова.
Хотя белок Ran управляет как импортом, так и экспортом белков в ядро, в каждом случае только активная форма (Ran:ГTФ) связывается с транспортёром. Форма Rаn:ГДФ) является неактивной и не может связаться с белком-транспортёром.
Некоторые белки могут перемещаться между цитозолем и нуклеоплазмой, экспонируя и маскируя свою нацеливающую последовательность.
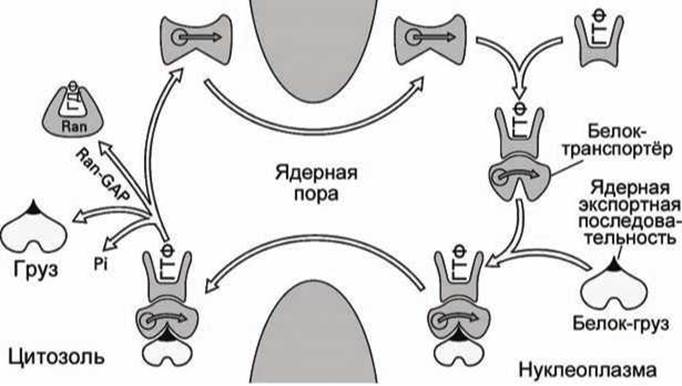
Рисунок 81 - Экспорт белков из ядра