Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Внутриклеточный транспорт белков
Трансмембранная транслокация
Транспорт белков в митохондрию. Митохондрия содержит собственную ДНК и синтезирует некоторые из необходимых для её функционирования белков, однако большинство митохондриальных белков кодируются ядерным геномом клетки. Такие белки синтезируются на рибосомах в цитозоле, а затем транспортируются в митохондрию. Как было отмечено выше, те белки, которые предназначены для матрикса митохондрии содержат на своём N-конце специфическую нацеливающую последовательность. Митохондриальные рецепторные белки распознают такую последовательность и соединяются с транслокационным комплексом, который разворачивает глобулу белка и пропускает полипептидную нить через обе мембраны митохондрии. После транслокации нацеливающая последовательность удаляется, и происходит фолдинг в нативную функциональную глобулу.
В процессах разворачивания белковой глобулы перед транслокацией и при вторичном фолдинге активно участвуют вспомогательные белки шапероны, которые предохраняют белковую нить от неспецифической агрегации.
Транспорт в пероксисомы. В большинство органелл, которые ограничены единственной мембраной, белки доставляются с помощью везикулярного транспорта (рисунок 76). Пероксисомы являются исключением из этого правила.
Белки пероксисом синтезируются в цитозоле, а затем транспортируются в пероксисомы. Пероксисомальные нацеливающие последовательности аминокислот в структуре этих белков связываются с белками- рецепторами импорта в пероксисомы ещё в цитозоле, и затем эти комплексы транспортируемых белков с рецепторами присоединяются к мембране пероксисомы, пересекают мембрану, в люмене пероксисомы транспортируемый белок высвобождается, а рецепторы переносятся обратно в цитозоль.
Синтез белка в шероховатом эндоплазматическом ретикулуме.
Наличие рибосом на мембранах шероховатого эндоплазматического ретикулума однозначно показывает, что это место синтеза белков. Однако в большинстве случаев на рибосомах шероховатого эндоплазматического синтезируются белки, не участвующие ни в каких процессах внутри данной клетки, "ненужные" ей, а иногда даже вредные для клетки.
Например, на рибосомах шероховатого эндоплазматического ретикулума клеток молочной железы в большом количестве синтезируется казеин молока, который совсем не нужен клеткам молочной железы, а на рибосомах шероховатого эндоплазматического ретикулума клеток пищеварительных желез синтезируются гидролитические ферменты, попадание которых в цитозоль неминуемо должно привести к самоперевариванию клетки и её гибели.
Однако этого не происходит, потому что синтезируемые белки переносятся через мембрану шероховатого эндоплазматического ретикулума в его полость (люмен) и тем самым сразу же изолируются от цитозоля и цитоплазматических структур.
Таким образом, роль шероховатого эндоплазматического ретикулума заключается не просто в синтезе белков на рибосомах его мембран, но и в изоляции этих белков от остальных цитозольных белков клетки.
Обобщённая схема биосинтеза белка представлена на рисунке 82.
Биосинтез белка (трансляция мРНК) всегда начинается в цитозоле (рисунок 82 (1)). Определённая последовательность из 15-60 аминокислот в начале цепи, обозначаемая как сигнальный пептид, указывает место синтеза. Если образующийся на рибосоме белок начинается с сигнального пептида (рисунок 82 (2)), ориентирующего белок на шероховатый эндоплазматический ретикулум, с ним связывается РНК-содержащая сигнал-узнающая частица, SRP (signal-recognition particle), и трансляция временно прерывается (рисунок 82 (3)).
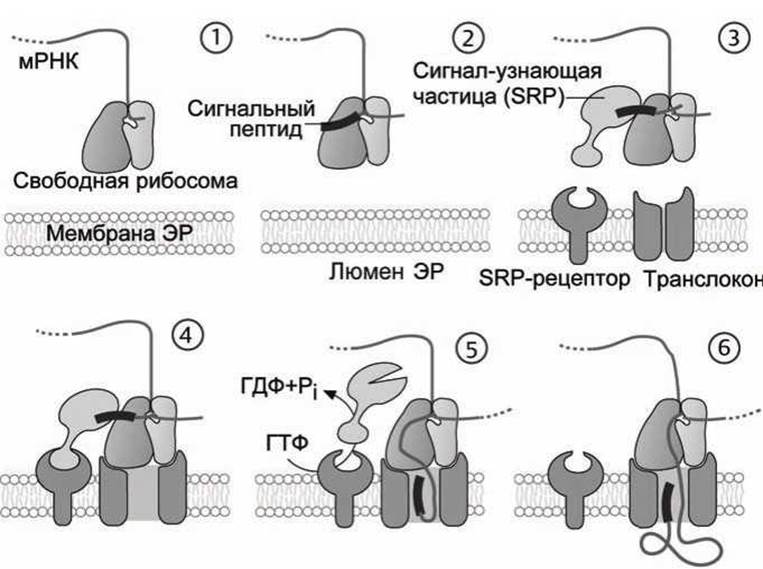
Рисунок 82 - Синтез белка в шероховатом эндоплазматическом ретикулуме
SRP связывает рибосому посредством SRP-рецептора с мембраной шероховатого эндоплазматического ретикулума (рисунок 82 (4)). Как только рибосома закрепится на мембране, SRP-частица диссоциирует от сигнального пептида и от SRP-рецептора (при этом гидролизуется ГТФ), и на рибосоме вновь начинается процесс трансляции (рисунок 82 (5)).
Белковая цепь на рибосоме растет и, ещё не свернувшись, проходит через мембрану по каналу, называемому транслоконом, в люмен шероховатого эндоплазматического ретикулума (рисунок 82 (6)).
После окончания трансляции сигнальный пептид секреторного белка отрезается специальным ферментом лидер-пептидазой, и белок уже не может покинуть полость шероховатого эндоплазматического ретикулума. Таким способом осуществляется направленное выведение (vectorial discharge) белков.
Прохождение растущего полипептида через мембрану может быть прервано соответствующим стоп-сигналом. В этом случае полипептид остаётся погружённым в мембрану и даёт начало интегральному мембранному белку. В ходе белкового синтеза возможно многократное прохождение растущей цепи через мембрану и возобновление синтеза вновь при посредстве сигнального пептида. Образующийся по такому механизму мембранный белок будет иметь множество трансмембранных участков.
Большинство белков, синтезированных на мембранах шероховатого эндоплазматического ретикулума, представляют собой гликопротеины, тогда как растворимые белки цитозоля не гликозилированы. Гликозилирование — это ещё одна важная биосинтетическая функция эндоплазматического ретикулума. Это гликозилирование оказывает влияние на дальнейшую судьбу белков.