Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Вторичный транспорт котранспортёрами
Nа+-симпортёры
Котранспортёры являются транспортными белками, которые переносят молекулы метаболитов (ионы и малые молекулы, такие как глюкоза или аминокислоты) против градиента концентрации, не используя энергию гидролиза АТФ. Для выполнения работы котранспортёры используют энергию, предварительно запасённую в электрохимическом градиенте ионов Na+ или Н+, для такого перемещения через мембрану молекул.
Например, энергетически выгодное движение ионов Na+ внутрь клетки, которое стимулируется и электрическим полем, и градиентом концентрации натрия, может быть сопряжено с перемещением транспортируемой молекулы, например, глюкозы, против её концентрационного градиента.
Важной особенностью такого совместного транспорта (или котранспорта) является то, что ни одна из молекул не может перемещаться через мембрану самостоятельно, возможно только совместное перемещение, поэтому такой транспорт называют сопряжённым.
Симпортом называется процесс переноса метаболитов белком-котранспортёром в том же направлении, в котором через мембрану переносятся ионы, градиент концентрации которых обеспечивает энергией работу мембранного белка (рисунок 52(6)). Если же перенос молекул и котранспортирующих ионов происходит во встречных направлениях, то такой процесс называется антипортом (рисунок 52(b)).
Некоторые котранспортёры переносят только катионы, другие - только анионы. Важным примером катионного котранспортёра является Na+/H+-aнтипортep, который выкачивает из клетки протоны, используя энергию потока ионов натрия внутрь клетки. Примером анионного котранспортёра является анионный белок антипортёр АЕ1, который осуществляет встречный перенос анионов Сl- и НСО-3 через мембрану.
Большинство клеток тканей животных импортируют глюкозу из крови с помощью транспортёров типа GLUT (рисунок 55), а направление трансмембранного переноса глюкозы задаётся градиентом концентрации.
Однако для некоторых клеток, таких как клетки, выстилающие слизистую тонкого кишечника или почечные протоки, необходимо импортировать глюкозу из люмена кишечника или протоков против значительного градиента концентрации. Такие клетки используют 2- Na+/1 -глгакоза-симпортёр - белок, который сопрягает импорт в клетку одной молекулы глюкозы с импортом двух ионов Na+
![]()
Свободная энергия для этого случая может быть записана в форме
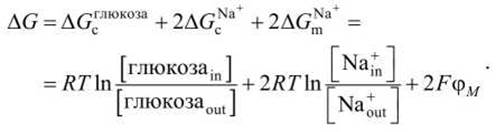
Здесь в ∆G входят как изменения свободной энергии за счёт градиента химического потенциала при переносе через мембрану 1 моля глюкозы (∆Gcглюкоза и 2 молей ионов натрия (2∆GсNa+), так и изменение свободной энергии за счёт градиента электрического потенциала при переносе 2 молей катионов натрия (2∆GNa+m). На рисунке 102 представлена схема формирования ∆G при переносе ионов натрия внутрь клетки.
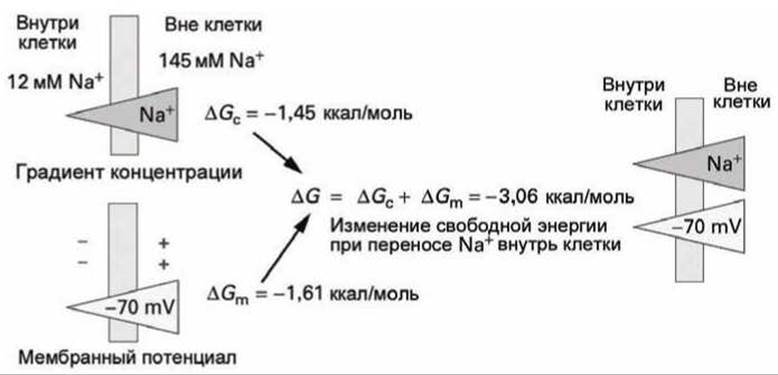
Рисунок 102 - Изменение свободной энергии при транспорте одного моля Na+ внутрь клетки
При транспорте двух молей ионов натрия через мембрану выигрыш в энергии составляет приблизительно 6 ккал/моль. При равновесии ∆G = 0, следовательно, равновесное соотношение концентраций глюкозы (определяемое из уравнения ![]() -6 ккал/моль) составляет
-6 ккал/моль) составляет ![]()
Поэтому перенос двух молей ионов натрия симпортёрами создаёт концентрацию глюкозы внутри клетки в 30 000 раз больше, чем вне клетки. Если бы симпортёр использовал энергию переноса только одного иона натрия, то соотношение концентраций глюкозы внутри и снаружи клетки было бы только 170.
Таким образом, сопряжение импорта глюкозы с переносом всего двух (а не одного) ионов натрия позволяет клетке аккумулировать почти в 200 раз больше глюкозы, чем это было бы возможно в случае 1-Na+/1-глюкоза транспортёра.
2-Nа+/1-глюкоза-симпортёр содержит 14 трансмембранных а-спиралей, и его транспортный цикл изображен на рисунке 103.
В отличие от, например, GLUT1-унипортёра (рисунок 55), конформационный переход в 2-Nа+/1-глюкоза-симпортёре происходит только когда все субстраты (два иона натрия и молекула глюкозы) присоединены к экзоплазматическому домену белка.
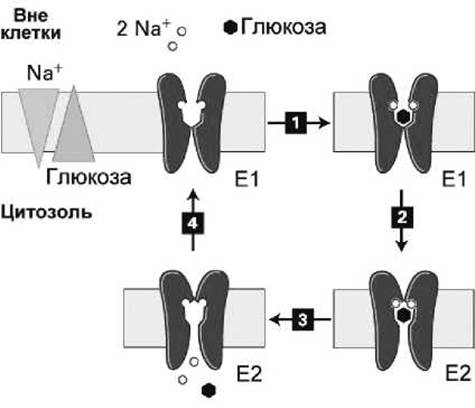
Рисунок 103 - Схема рабочего цикла 2-Nа+/1-глюкоза-симпортёра