Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Вторичный транспорт котранспортёрами
Трансэпителиальный транспорт
Соединения между клетками в составе тканей и органов многоклеточных организмов образуются специализированными структурами, которые называются межклеточными контактами. В простейшем случае межклеточные соединения (которые часто также называют контактами или переходами) разделяют на плотные соединения (tight junctions) и щелевые соединения (gap junction, см п. 16.1).
Плотные контакты герметизируют межклеточные соединения в эпителиях и не позволяют метаболитам проникать сквозь эпителий, минуя клетки эпителия (рисунок 109(a)).
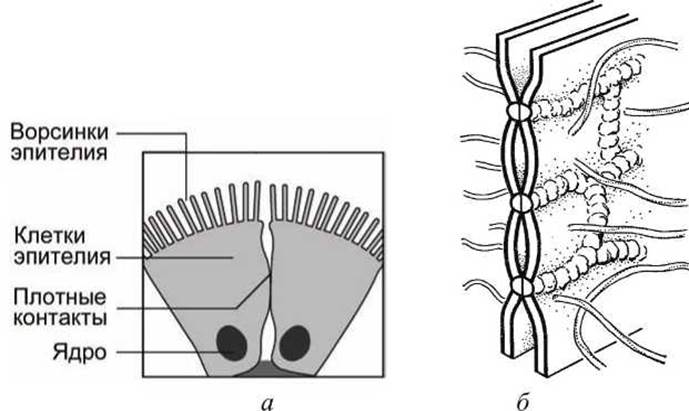
Рисунок 109 - Плотные межклеточные контакты: а - контакт двух клеток эпителия, б - строение плотного контакта
Плотные контакты разделяются на зону залипання и зону слипания.
В зоне залипания мембраны соседних клеток сливаются своими наружными слоями, эта зона непроницаема для макромолекул и ионов.
В зоне слипания мембраны разделены щелью 10-20 нм.
Щелевые контакты (будут рассмотрены подробнее ниже в п. 16.1) соединяют цитозоли клеток и позволяют ионам и малым молекулами переходить из клетки в клетку.
Через эпителий возможны два пути транспорта метаболитов: трансклеточный и параклеточный (рисунок 110).
Основным трансэпителиальным транспортом является трансклеточный путь. Параклеточный путь, по которому могут перемещаться некоторые ионы и малые молекулы, является дополнительным и зависит от конкретного белкового состава межклеточных контактов и от физиологического состояния эпителиальных клеток.
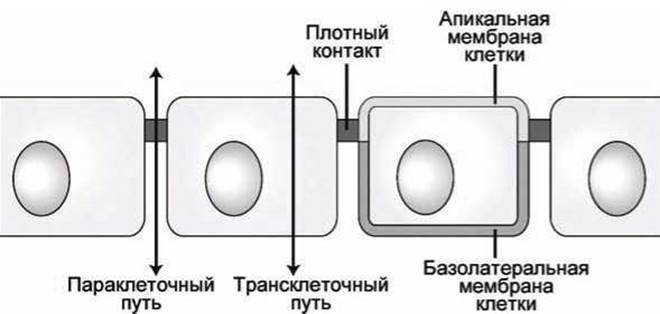
Рисунок 110 - Трансклеточный и параклеточный пути трансэпителиального транспорта
В случае кишечного эпителия поглощение питательных веществ из люмена кишечника происходит с люминальной стороны через апикальную мембрану клеток эпителия, а затем происходит экспорт этих веществ в кроветoк с серозной стороны (serosal side) через базолатеральную мембрану (рисунки 110 и 19).
Такой двухстадийный процесс называется трансклеточным транспортом.
Клетки эпителия кишечника (также как и клетки любого другого эпителия) называют поляризованными, поскольку апикальные и базолатеральные домены плазматической мембраны содержат различные наборы мембранных белков. Эти два домена плазматической мембраны разделены плотными межклеточными соединениями.
Апикальная часть плазматической мембраны, которая контактирует с люменом кишечника, специализируется на адсорбции из люмена сахаров, аминокислот и других молекул, которые образуются в люмене в процессе пищеварения пищеварительными ферментами. Площадь поверхности апикальной мембраны значительно увеличена за счёт формирования на ней микроворсинок, что позволяет интенсифицировать процесс адсорбции метаболитов.
На рисунке 111 показаны мембранные белки, которые осуществляют адсорбцию глюкозы из люмена кишечника в кровь.
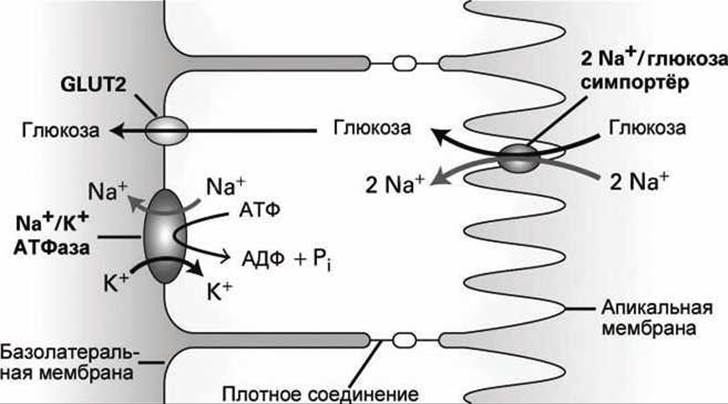
Рисунок 111 - Трансклеточный транспорт глюкозы из люмена кишечника в кровь
На первом этапе этого процесса 2-Na+/1-глюкоза-симпортёры, расположенные на мембранах микроворсинок, импортируют из люмена кишечника молекулы глюкозы против градиента их концентрации через апикальную поверхность эпителиальной клетки. Такой симпортёр сопрягает энергетически невыгодный транспорт глюкозы внутрь клетки с энергетически выгодным процессом переноса двух ионов натрия внутрь клетки.
В стационарном состоянии все ионы натрия, переносимые внутрь клетки в процессах симпорта Na+/глюкоза или Na+/аминокислоты, выкачиваются через базолатеральную мембрану Na+/К+-ЛТФазами, которые обнаружены только в базолатеральных мембранах клеток кишечного эпителия. В результате концентрация ионов Na+ в цитозоле этих клеток поддерживается на низком уровне. И можно сделать вывод, что источником энергии для первого этапа трансклеточного транспорта глюкозы служит именно гидролиз АТФ Na+/К+-АТФазами.
На втором этапе молекулы глюкозы (и аминокислот), которые были сконцентрированы на первом этапе в цитозоле клеток эпителия, перемещаются под действием концентрационного градиента через базолатеральную мембрану в кровь с помощью мембранных белков- унипортёров. В случае транспорта глюкозы таким унипортёром является белок GLUT2 (см. п. 6.5).
Суммарным результатом этих двух этапов является транспорт ионов Na+, глюкозы и аминокислот из люмена кишечника через кишечный эпителий во внеклеточную среду, которая окружает базолатеральную поверхность клеток кишечного эпителия. Плотные соединения предохраняют эти молекулы от обратной диффузии в люмен кишечника.
Увеличение осмоса вследствие трансклеточного транспорта солей, сахаров и аминокислот приводит к интенсификации осмотического процесса переноса молекул воды из люмена кишечника в кровь. Поэтому, в определённом смысле, транспортируемые молекулы метаболитов "прихватывают" с собой молекулы воды.
Наличие такого "сопровождающего" потока воды используют в медицинских целях в ситуациях сильного обезвоживания (дегидрации) организма (при дизентерии, холере и других кишечных инфекциях, приводящих к высокой смертности, особенно у детей). В этих случаях дают больным пить водный раствор сахара и соли (вместе, но не по отдельности). В результате осмотического потока воды из люмена кишечника в кровь происходит частичная регидрация организма, что облегчает состояние пациента и позволяет проводить длительную терапию.