Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Потенциал действия
Свойства потенциала действия
Электрические нервные импульсы (потенциалы действия) в живом организме передают информацию от рецепторов к нейронам мозга и от нейронов мозга к мышцам. Живой организм является электрифицированной системой, и без электричества нет жизни.
Потенциал действия был открыт раньше потенциала покоя. Животное электричество известно давно. Разряды электрического угря (происходящие при напряжении до 600 В, с током около 60 А и длительностью порядка миллисекунды) использовались медициной ещё в Древнем Риме для лечения подагры, головной боли, эпилепсии. Электрический нервный импульс открыл Луиджи Гальвани, профессор анатомии в г. Болонья. Результаты его электрофизиологических опытов изложены в книге "Трактат о силах электричества при мышечном движении" (1791 г.).
Гальвани открыл, что мышечные сокращения конечностей препарированной лягушки могут вызываться электрическим импульсом, и что сама живая система является источником электрического импульса. Открытие Гальвани сыграло выдающуюся роль в развитии физики, электротехники, электрохимии, физиологии, биофизики, медицины.
Однако огромная популярность идей Гальвани привела к их профанации (гальванизация трупов, гальванизм прикосновений и взглядов и т.д.), что вызывало недоверие ученых к экспериментам Гальвани. Младший современник Гальвани профессор физики Алессандро Вольта был яростным противником идеи животного электричества (за исключением особых случаев электрических рыб: электрического угря и электрического ската). В своих экспериментах он исключил биологический объект и показал, что электрический ток может быть получен при контакте набора металлов, разделённых электролитом (вольтов столб). Так был открыт химический источник тока (названный, однако, позже в честь его научного противника гальваническим элементом).
В XIX веке утвердилось примитивное представление о распространении электрических токов по нервам, как по проводам. Однако Гельмгольцем во второй половине XIX века было показано, что скорость распространения нервного импульса составляет лишь 1-100 м/с, это значительно меньше, чем скорость распространения электрического импульса по проводам (3 ∙ 108м/с). Поэтому к концу XIX века гипотеза электрической природы нервного импульса была отвергнута большинством физиологов. Было выдвинуто предположение о распространении по нервным волокнам химической реакции.
Позднее было показано, что медленное распространение электрического нервного импульса связано с медленной перезарядкой конденсаторов, которые представляют собой клеточные мембраны, через большие сопротивления. Постоянная времени перезарядки мембраны т = RC велика, так как велики ёмкость мембраны С и сопротивление R нервного волокна.
То, что нервный импульс представляет собой импульс электрического тока, было доказано лишь к середине 20-го века, в основном, в работах английского физиолога Алана Ходжкина и его сотрудников. В 1963 году А.Л. Ходжкину (Alan Lloyd Hodgkin), Э.Ф. Хаксли (Andrew Fielding Huxley) и Дж.К. Эклсу (John Carew Eccles) была присуждена Нобелевская премия по физиологии и медицине "за открытия, касающиеся ионных механизмов возбуждения и торможения в периферических и центральных участках нервных клеток".
Потенциалом действия (action potential) называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения.
Опыты по исследованию потенциала действия проведены (в основном Ходжкиным и его сотрудниками) на гигантских аксонах кальмара методом микроэлектродов с использованием высокоомных измерителей напряжения, а также методом меченых атомов.
В опытах по исследованию потенциала действия использовали два микроэлектрода, введённых в аксон (рисунок 113).
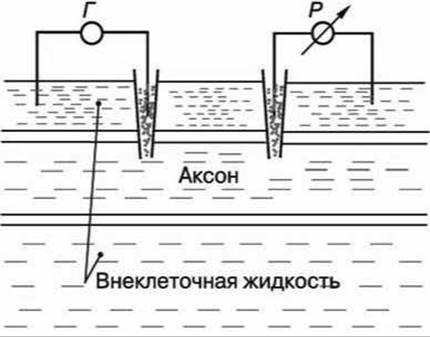
Рисунок 113 - Схема опыта по исследованию потенциала действия
На первый микроэлектрод подаётся импульс с амплитудой V от генератора Г прямоугольных импульсов, меняющий мембранный потенциал. Мембранный потенциал измеряется при помощи второго микроэлектрода высокоомным регистратором напряжения Р.
Отрицательный возбуждающий импульс вызывает смещение мембранного потенциала лишь на короткое время, спустя которое мембранный потенциал быстро пропадает, и восстанавливается потенциал покоя (рисунок 114). Увеличение амплитуды отрицательного возбуждающего импульса сопровождается гиперполяризацией мембраны.
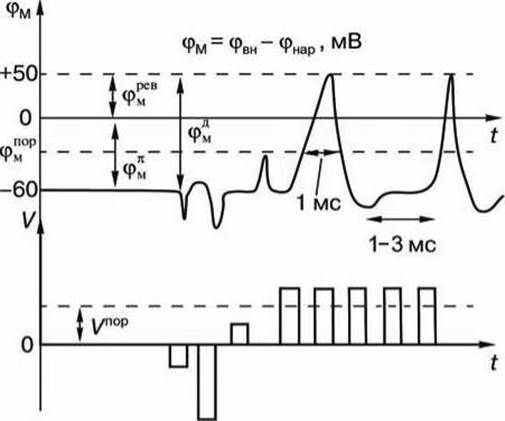
Рисунок 114 - Потенциал действия: V - амплитуда прямоугольного импульса; Vпоp - пороговое значение амплитуды; t - время, φМ - мембранный потенциал; φвн, фнар - потенциалы на внутренней и наружной поверхностях мембраны; φМД - амплитуда потенциала действия; φМ - уровень потенциала покоя; φМпор - пороговое значение мембранного потенциала; φМрев - мембранный потенциал реверсии
Потенциал действия не формируется также и в том случае, когда возбуждающий импульс положительный (деполяризующий), но его амплитуда меньше порогового значения Vпор.
Однако, если амплитуда положительного, деполяризующего импульса окажется больше значения Vпор, φм становится больше φпорМ и в мембране развивается процесс, в результате которого происходит резкое повышение мембранного потенциала, и мембранный потенциал φм даже меняет свой знак - становится положительным (φвн > φнар) - происходит деполяризация мембраны.
Достигнув некоторого положительного значения φревМ - потенциала реверсии, мембранный потенциал возвращается к значению потенциала покоя φПМ, совершив нечто вроде затухающего колебания. В нервных волокнах и скелетных мышцах длительность потенциала действия около 1 мс (а в сердечной мышце около 300 мс).
После снятия возбуждения ещё в течение 1—3 мс в мембране нейрона наблюдаются некоторые остаточные явления, во время которых мембрана рефрактерна (невозбудима).
Новый деполяризующий потенциал V > Vпop может вызвать образование нового потенциала действия только после полного возвращения мембраны в состояние покоя. Причём амплитуда потенциала действия φДМ = |φПМ | + |φревМ| не зависит от амплитуды деполяризующего потенциала (если только V > Vпop).
Если в покое мембрана поляризована (потенциал цитоплазмы отрицателен по отношению к внеклеточной среде), то при возбуждении происходит деполяризация мембраны (потенциал внутри клетки положителен), и после снятия возбуждения происходит реполяризация мембраны.
Характерные свойства потенциала действия:
1) наличие порогового значения деполяризующего потенциала;
2) закон "всё или ничего", то есть, если деполяризующий потенциал больше порогового, развивается потенциал действия, амплитуда которого не зависит от амплитуды возбуждающего импульса, и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой;
3) существует период рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения;
4) в момент возбуждения резко уменьшается сопротивление мембраны (у аксона кальмара сопротивление мембраны уменьшается в 40 раз от 0,1 Ом∙м2 в покое до 0,0025 Ом∙м2 при возбуждении).